





И активентранспорт. Пасивният транспорт се извършва без консумация на енергия по електрохимичен градиент. Пасивните включват дифузия (проста и улеснена), осмоза, филтрация. Активният транспорт изисква енергия и протича срещу концентрация или електрически градиенти.
Активен транспорт
Това е транспортирането на вещества в противоречие с концентрацията или електрическите градиенти, което се случва с разхода на енергия. Прави се разлика между първичен активен транспорт, който изисква енергия от АТФ, и вторичен (за сметка на АТФ, създаване на градиенти на йонна концентрация от двете страни на мембраната и енергията на тези градиенти се използва за транспорт).
Първичният активен транспорт се използва широко в тялото. Той участва в създаването на електрическа потенциална разлика между вътрешната и външната страна на клетъчната мембрана. С помощта на активен транспорт се създават различни концентрации на Na +, K +, H +, SI "" и други йони в средата на клетката и в извънклетъчната течност.
Транспортът на Na+ и K+ е по-добре проучен - Na+, -K + -Hacoc. Този транспорт се осъществява с участието на глобуларен протеин с молекулно тегло около 100 000. Протеинът има три Na + свързващи места на вътрешната повърхност и две K + свързващи места на външната повърхност. На вътрешната повърхност на протеина се наблюдава висока АТФазна активност. Енергията, генерирана по време на хидролизата на АТФ, води до конформационни промени в протеина и в същото време три Na + йона се отстраняват от клетката и в нея се въвеждат два K + йона.С помощта на такава помпа, a висока концентрация на Na + се създава в извънклетъчната течност и висока концентрация на K + - в клетъчната течност.
Напоследък интензивно се изследват Ca2 + помпи, благодарение на които концентрацията на Ca2 + в клетката е десетки хиляди пъти по-ниска, отколкото извън нея. Са2+ помпи има в клетъчната мембрана и в клетъчните органели (саркоплазмен ретикулум, митохондрии). Ca2+ помпите също функционират благодарение на протеин носител в мембраните. Този протеин има висока АТФазна активност.
Вторичен активен транспорт. Благодарение на първичния активен транспорт се създава висока концентрация на Na + извън клетката, възникват условия за дифузия на Na + в клетката, но заедно с Na + други вещества могат да влязат в нея. Този транспорт е насочен в една посока и се нарича симпорт. В противен случай влизането на Na + стимулира излизането на друго вещество от клетката, това са два потока, насочени в различни посоки - антипорт.
Пример за симпорт би бил транспортирането на глюкоза или аминокиселини заедно с Na+. Протеинът носител има две места за свързване с Na + и за свързване с глюкоза или аминокиселина. Идентифицирани са пет различни протеина, които свързват пет вида аминокиселини. Известни са и други видове симпорт - транспорт на N + заедно с в клетката, K + и Cl- от клетката и др.
В почти всички клетки има антипортов механизъм - Na + отива в клетката и Ca2 + я напуска или Na + отива в клетката и H + излиза от нея.
Mg2 +, Fe2 +, HCO3- и много други вещества се транспортират активно през мембраната.
Пиноцитозата е един от видовете активен транспорт. Това се дължи на факта, че някои макромолекули (главно протеини, макромолекулите на които имат диаметър 100-200 nm) се прикрепят към мембранните рецептори. Тези рецептори са специфични за различни протеини. Прикрепването им е съпроводено с активиране на съкратителните протеини на клетката - актин и миозин, които образуват и затварят кухина с този извънклетъчен протеин и малко количество извънклетъчна течност. В този случай се образува пиноцитозна везикула. Той секретира ензими, които хидролизират този протеин. Продуктите от хидролизата се абсорбират от клетките. Пиноцитозата изисква АТФ енергия и наличието на Ca2+ в извънклетъчната среда.
По този начин има много видове транспорт на вещества през клетъчните мембрани. Различни видове транспорт могат да възникнат от различни страни на клетката (в апикалната, базалната, латералната мембрана). Пример за това са процеси, протичащи в
12345Напред ⇒
Бележки от лекции №3.
Предмет. Субклетъчно и клетъчно ниво на организация на живия живот.
Структурата на биологичните мембрани.
Основата на биологичната мембрана на всички живи организми е двойна фосфолипидна структура. Фосфолипидите на клетъчните мембрани са триглицериди, в които една от мастните киселини е заменена с фосфорна киселина. Хидрофилните "глави" и хидрофобните "опашки" на фосфолипидните молекули са ориентирани така, че се появяват два реда молекули, чиито глави покриват "опашките" от вода.
В тази фосфолипидна структура са интегрирани протеини с различни размери и форми.
Индивидуалните свойства и характеристики на мембраната се определят главно от протеини. Различният протеинов състав определя разликата в структурата и функциите на органелите на всеки животински вид. Влиянието на състава на мембранните липиди върху техните свойства е много по-слабо.
Транспорт на вещества през биологични мембрани.
![]()
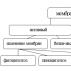
Транспортът на веществата през мембраната се разделя на пасивен (без разход на енергия по концентрационен градиент) и активен (с разход на енергия).
Пасивен транспорт: дифузия, улеснена дифузия, осмоза.
Дифузията е движението на частици, разтворени в среда, от зона с висока концентрация към зона с ниска концентрация (разтваряне на захар във вода).
Улеснената дифузия е дифузия, използваща канален протеин (влизане на глюкоза в червените кръвни клетки).
Осмозата е движението на частици на разтворителя от област с по-ниска концентрация на разтворено вещество към област с висока концентрация (червеното кръвно телце набъбва и се спуква в дестилирана вода).
Активният транспорт се разделя на транспорт, свързан с промени във формата на мембраната и транспорт чрез протеини с ензимна помпа.
От своя страна транспортът, свързан с промени във формата на мембраната, се разделя на три типа.
Фагоцитозата е улавянето на плътен субстрат (левкоцит-макрофаг улавя бактерия).
Пиноцитозата е улавяне на течности (хранене на ембрионални клетки в първите етапи на вътрематочно развитие).
Транспортирането чрез помпени ензимни протеини е движението на вещество през мембрана с помощта на протеини носители, интегрирани в мембраната (транспорт на натриеви и калиеви йони съответно „навън“ и „вътре“ в клетката).
По посока транспортът се разделя на екзоцитоза(от клетката) и ендоцитоза(в клетка).
Класификация на клетъчните компонентиизвършвани по различни критерии.
Въз основа на наличието на биологични мембрани органелите се делят на двойномембранни, едномембранни и немембранни.
Според функциите си органелите се делят на неспецифични (универсални) и специфични (специализирани).
В случай на повреда те се класифицират на жизненоважни и възстановими.
Според принадлежността към различни групи живи същества: растения и животни.
Мембранните (едно- и двумембранни) органели имат подобна структура от химична гледна точка.
Двойни мембранни органели.
Ядро. Ако клетките на даден организъм имат ядро, тогава те се наричат еукариоти. Ядрената обвивка има две близко разположени мембрани. Между тях е перинуклеарното пространство. Ядрената мембрана има дупки, наречени пори. Нуклеолите са частите от ядрото, отговорни за синтеза на РНК. В ядрата на някои клетки на жените обикновено се секретира 1 телце на Barr - неактивна Х хромозома. Когато ядрото се раздели, всички хромозоми стават видими. Извън деленето хромозомите обикновено не се виждат. Ядреният сок е кариоплазмата. Ядрото осигурява съхранението и функционирането на генетичната информация.
Митохондриите. Вътрешната мембрана има кристи, които увеличават вътрешната повърхност за ензимите на аеробното окисление. Митохондриите имат собствена ДНК, РНК и рибозоми. Основната функция е завършването на окислението и фосфорилирането на ADP
ADP+P=ATP.
Пластиди (хлоропласти, хромопласти, левкопласти). Пластидите имат свои собствени нуклеинови киселини и рибозоми. Стромата на хлоропластите съдържа дисковидни мембрани, събрани в купчини, където се намира хлорофилът, отговорен за фотосинтезата.
Хромопластите имат пигменти, които определят жълтия, червения, оранжевия цвят на листата, цветята и плодовете.
Левкопластите съхраняват хранителни вещества.
Едномембранни органели.
Външната цитоплазмена мембрана отделя клетката от външната среда. Мембраната има протеини, които изпълняват различни функции. Има рецепторни протеини, ензимни протеини, помпани протеини и канални протеини. Външната мембрана има селективна пропускливост, позволяваща транспортирането на вещества през мембраната.
Някои мембрани съдържат елементи от надмембранния комплекс - клетъчната стена при растенията, гликокаликса и микровилите на чревните епителни клетки при хората.
Има апарат за контакт със съседни клетки (например десмозоми) и подмембранен комплекс (фибриларни структури), който осигурява стабилността и формата на мембраната.
Ендоплазменият ретикулум (ER) е система от мембрани, които образуват цистерни и канали за взаимодействия в клетката.
Има гранулиран (груб) и гладък EPS.
Гранулираният ER съдържа рибозоми, където се осъществява биосинтезата на протеини.
На гладкия ER се синтезират липиди и въглехидрати, глюкозата се окислява (стадий без кислород), ендогенни и екзогенни (чужди ксенобиотици, включително лекарствени) вещества се неутрализират. За неутрализация гладкият EPS съдържа ензимни протеини, които катализират 4 основни типа химични реакции: окисление, редукция, хидролиза, синтез (метилиране, ацетилиране, сулфатиране, глюкурониране). В сътрудничество с апарата на Голджи, ER участва в образуването на лизозоми, вакуоли и други едномембранни органели.
Апаратът на Голджи (ламеларен комплекс) е компактна система от плоски мембранни цистерни, дискове и везикули, която е тясно свързана с ER. Ламеларният комплекс участва в образуването на мембрани (например за лизозоми и секреторни гранули), които отделят хидролитичните ензими и други вещества от клетъчното съдържание.
Лизозомите са везикули, съдържащи хидролитични ензими. Лизозомите участват активно във вътреклетъчното храносмилане и фагоцитоза. Те усвояват обекти, уловени от клетката, сливайки се с пиноцитни и фагоцитни везикули. Те могат да усвояват собствените си износени органели. Фаговите лизозоми осигуряват имунна защита. Лизозомите са опасни, защото при разрушаване на обвивката им може да настъпи автолиза (самосмилане) на клетката.
Пероксизомите са малки, едномембранни органели, съдържащи ензима каталаза, който неутрализира водородния пероксид. Пероксизомите са органели, които предпазват мембраните от пероксидация на свободните радикали.
Вакуолите са едномембранни органели, характерни за растителните клетки. Техните функции са свързани с поддържане на тургора и (или) съхраняване на вещества.
Немембранни органели.
Рибозомите са рибонуклеопротеини, състоящи се от големи и малки рРНК субединици. Рибозомите са мястото на сглобяване на протеини.
Фибриларните (нишковидни) структури са микротубули, междинни нишки и микрофиламенти.
Микротубули. Структурата наподобява мъниста, чиято нишка е навита в плътна пружинна спирала. Всяко "мънисто" представлява тубулин протеин. Диаметърът на тръбата е 24 nm. Микротубулите са част от система от канали, които осигуряват вътреклетъчния транспорт на вещества. Те укрепват цитоскелета, участват в образуването на вретеното, центриолите на клетъчния център, базалните тела, ресничките и флагелите.
Клетъчният център е част от цитоплазмата с две центриоли, образувани от 9 триплета (по 3 микротубули). Така всеки центриол се състои от 27 микротубули. Смята се, че клетъчният център е в основата на образуването на нишките на вретеното на клетъчното делене.
Базалните тела са основите на ресничките и флагелите. В напречно сечение ресничките и камшичетата имат девет чифта микротубули около обиколката и една двойка в центъра, за общо 18 + 2 = 20 микротубули. Ресничките и флагелите осигуряват движението на микроорганизми и клетки (сперматозоиди) в тяхното местообитание.
Междинните нишки имат диаметър 8-10 nm. Те осигуряват функциите на цитоскелета.
Микрофиламентите с диаметър 5-7 nm са съставени предимно от протеина актин. Във взаимодействие с миозина те са отговорни не само за мускулните контракции, но и за контрактилната активност на немускулните клетки. По този начин промените във формата на мембраната по време на фагоцитоза и активността на микровилите се обясняват с работата на микрофиламентите.
Включванията са натрупвания на вещества в клетката, които не са ограничени от вътреклетъчни мембрани (капки мазнини, бучки гликоген).
Разделянето на органелите на неспецифични (универсални) и специфични (специализирани) е доста произволно. Органелите със специално предназначение включват реснички и флагели, микровили и мускулни микрофиламенти.
Животинските клетки се различават от растителните по липсата на целулоза и клетъчна стена, вакуоли с клетъчен сок и пластиди. Растителните клетки на висшите растения нямат реснички или флагели. Растенията нямат центриоли.
Ако ядрото и митохондриите са повредени (цианидно отравяне), клетъчната смърт е неизбежна, тъй като информацията и енергията са блокирани. Ядрото и митохондриите се считат за жизненоважни органели. Когато други органели са унищожени, съществува фундаментална възможност за тяхното възстановяване.
12345Напред ⇒
Свързана информация:
Търсене в сайта:
Биологични мембрани(лат. мембрана мембрана, мембрана) - функционално активни повърхностни структури с дебелина няколко молекулярни слоя, ограничаващи цитоплазмата и повечето органели на клетката, а също така образувайки единна вътреклетъчна система от тубули, гънки и затворени зони.
Биологичните мембрани присъстват във всички клетки. Тяхното значение се определя от важността на функциите, които изпълняват в процеса на нормалния живот, както и от разнообразието от заболявания и патологични състояния, които възникват от различни нарушения на мембранните функции и се проявяват на почти всички нива на организация - от клетъчните и субклетъчните системи към тъканите, органите и тялото като цяло.
Мембранните структури на клетката са представени от повърхностни (клетъчни или плазмени) и вътреклетъчни (субклетъчни) мембрани. Името на вътреклетъчните (субклетъчните) мембрани обикновено зависи от името на структурите, които съдържат или образуват. По този начин има митохондриални, ядрени, лизозомни мембрани, мембрани на ламеларния комплекс на апарата на Голджи, ендоплазмен ретикулум, саркоплазмен ретикулум и др. (вж. клетка). Дебелина на биологичните мембрани - 7-10 nm, но общата им площ е много голяма, например в черния дроб на плъх тя е няколкостотин квадратни метра.
Химичен състав и структура на биологичните мембрани.Съставът на биологичните мембрани зависи от техния вид и функция, но основните компоненти са липидиИ протеини,и въглехидрати(малка, но изключително важна част) и вода (повече от 20% от общото тегло).
Липиди. В биологичните мембрани се намират три класа липиди: фосфолипиди, гликолипиди и стероиди. В мембраните на животинските клетки повече от 50% от всички липиди са фосфолипиди - глицерофосфолипиди (фосфатидилхолин, фосфатидилетаноламин, фосфатидилсерин, фосфатидилинозитол) и сфингофосфолипиди (керамидни производни, сфингомиелин). Гликолипидите са представени от цереброзиди, сулфатиди и ганглиозиди, а стероидите са главно холестерол (около 30%). Липидните компоненти на биологичните мембрани съдържат различни мастни киселини, но палмитинова, олеинова и стеаринова киселина преобладават в мембраните на животински клетки. Фосфолипидите играят основната структурна роля в биологичните мембрани. Имат изразена способност да образуват двуслойни структури (двуслойни) при смесване с вода, което се дължи на химичната структура на фосфолипидите, чиито молекули се състоят от хидрофилна част - "глава" (остатък от фосфорна киселина и полярна група, прикрепена към него, например холин) и хидрофобна част - "опашка" (обикновено две вериги на мастни киселини). Във водна среда фосфолипидите на двуслоя са разположени по такъв начин, че остатъците от мастни киселини са обърнати към вътрешността на двуслоя и следователно са изолирани от околната среда, а хидрофилните „глави“, напротив, са обърнати навън . Липидният двоен слой е динамична структура: липидите, които го образуват, могат да се въртят, да се движат странично и дори да се преместват от слой на слой (преход на тригер). Тази структура на липидния двоен слой формира основата на съвременните идеи за структурата на биологичните мембрани и определя някои важни свойства на биологичните мембрани, например способността да служат като бариера и да не позволяват на молекулите на веществата, разтворени във вода, да преминат през ( ориз .). Нарушаването на двуслойната структура може да доведе до нарушаване на бариерната функция на мембраните.
Холестеролът в биологичните мембрани играе ролята на двуслоен модификатор, придавайки му известна твърдост чрез увеличаване на плътността на „опаковането“ на фосфолипидните молекули.
Гликолипидите имат различни функции: те са отговорни за приемането на определени биологично активни вещества, участват в тъканната диференциация и определят видовата специфичност.
катерицибиологичните мембрани са изключително разнообразни. Тяхното молекулно тегло е предимно 25 000 - 230 000.
Протеините могат да взаимодействат с липидния двоен слой поради електростатични и/или междумолекулни сили. Те могат да бъдат сравнително лесно отстранени от мембраната. Този тип протеин включва цитохром с (молекулно тегло около 13 000), който се намира на външната повърхност на вътрешната мембрана на митохондриите.
Тези протеини се наричат периферни или външни. Други протеини, наречени интегрални или вътрешни, се характеризират с факта, че една или повече полипептидни вериги са вградени в двойни слоеве или ги пресичат, понякога повече от веднъж (например гликофорин, АТФ транспортази, бактериородопсин). Частта от протеина, която е в контакт с хидрофобната част на липидния бислой, има спирална структура и се състои от неполярни аминокиселини, поради което възниква хидрофобно взаимодействие между тези компоненти на протеините и липидите. Полярните групи от хидрофилни аминокиселини директно взаимодействат с близките мембранни слоеве, както от едната, така и от другата страна на двойния слой. Протеиновите молекули, подобно на липидните молекули, са в динамично състояние; те също се характеризират с ротационна, странична и вертикална подвижност. Това е отражение не само на тяхната собствена структура, но и на функционална дейност. което до голяма степен се определя от вискозитета на липидния бислой, който от своя страна зависи от състава на липидите, относителното съдържание и вида на вериги на ненаситени мастни киселини. Това обяснява тесния температурен диапазон на функционалната активност на мембранно свързаните протеини.
Мембранните протеини изпълняват три основни функции: каталитична (ензими), рецепторна и структурна. Подобно разграничение обаче е доста произволно и в някои случаи един и същ протеин може да изпълнява както рецепторни, така и ензимни функции (например инсулин).
Брой на мембраната ензимив клетката е доста голям, но разпределението им в различните видове биологични мембрани не е еднакво. Някои ензими (маркери) присъстват само в мембрани от определен тип (например Na, K-ATPase, 5-нуклеотидаза, аденилат циклаза - в плазмената мембрана; цитохром Р-450, NADPH дехидрогеназа, цитохром b5 - в мембраните на ендоплазмения ретикулум; моноаминооксидаза - във външната мембрана на митохондриите и цитохром С оксидаза, сукцинат дехидрогеназа - във вътрешната мембрана; кисела фосфатаза - в мембраната на лизозомите).
Рецепторните протеини, специфично свързващи нискомолекулни вещества (много хормони, медиатори), обратимо променят формата си. Тези промени предизвикват химични реакции вътре в клетката. По този начин клетката получава различни сигнали, идващи от външната среда.
Структурните протеини включват цитоскелетни протеини, съседни на цитоплазмената страна на клетъчната мембрана. В комбинация с микротубулите и микрофиламентите на цитоскелета, те осигуряват на клетката устойчивост на промени в нейния обем и създават еластичност. Тази група включва и редица мембранни протеини, чиито функции не са установени.
Въглехидратив биологичните мембрани те се комбинират с протеини (гликопротеини) и липиди (гликолипиди). Въглехидратните вериги на протеините са олиго- или полизахаридни структури, съдържащи глюкоза, галактоза, невраминова киселина, фукоза и маноза. Въглехидратните компоненти на биологичните мембрани се отварят главно в извънклетъчната среда, образувайки на повърхността на клетъчните мембрани много разклонени структури, които са фрагменти от гликолипиди или гликопротеини. Техните функции са свързани с контрол на междуклетъчното взаимодействие, поддържане на имунния статус на клетката и осигуряване на стабилността на протеиновите молекули в биологичните мембрани. Много рецепторни протеини съдържат въглехидратни компоненти. Пример са антигенните детерминанти на кръвните групи, представени от гликолипиди и гликопротеини.
Функции на биологичните мембрани.Бариерна функция. За клетките и субклетъчните частици биологичните мембрани служат като механична бариера, която ги отделя от външното пространство. Функционирането на клетката често се свързва с наличието на значителни механични градиенти на нейната повърхност, главно поради осмотично и хидростатично налягане. Основното натоварване в този случай се поема от клетъчната стена, чиито основни структурни елементи при висшите растения са целулоза, пектин и екстепин, а при бактериите - муреин (сложен полизахарид-пептид). В животинските клетки няма нужда от твърда мембрана. Известна твърдост се дава на тези клетки от специални протеинови структури на цитоплазмата, съседни на вътрешната повърхност на плазмената мембрана.
Трансфер на веществачрез биологични мембрани се свързва с такива важни биологични явления като вътреклетъчна йонна хомеостаза, биоелектрични потенциали, възбуждане и провеждане на нервни импулси, съхранение и трансформация на енергия и др. (см. Биоенергия). Има пасивен и активен транспорт (пренос) на неутрални молекули, вода и йони през биологични мембрани. Пасивният транспорт не е свързан с разход на енергия, той се осъществява чрез дифузия по концентрационни, електрически или хидростатични градиенти. Активният транспорт протича срещу градиенти, свързан е с разхода на енергия (главно енергията на хидролизата на АТФ) и е свързан с работата на специализирани мембранни системи (мембранни помпи). Има няколко вида транспорт. Ако дадено вещество се транспортира през мембрана, независимо от наличието и преноса на други съединения, тогава този тип транспорт се нарича еднопортов. Ако транспортът на едно вещество е свързан с транспорта на друго, тогава говорим за котранспорт, с еднопосочен транспорт, наречен симпорт, и противоположно насочен транспорт, наречен антипорт. Специална група включва пренос на вещества чрез екзо- и пиноцитоза.
Пасивният трансфер може да се осъществи чрез проста дифузия през липидните двойни слоеве на мембраната, както и чрез специализирани образувания - канали. Чрез дифузия през мембраната в клетката проникват незаредени молекули, силно разтворими в липиди, вкл. много отрови и лекарства, както и кислород и въглероден диоксид. Каналите са липопротеинови структури, които обхващат мембраните. Те служат за транспортиране на определени йони и могат да бъдат в отворено или затворено състояние. Проводимостта на канала зависи от мембранния потенциал, който играе важна роля в механизма на генериране и провеждане на нервните импулси.
В някои случаи преносът на материя съвпада с посоката на градиента, но значително надвишава скоростта на простата дифузия. Този процес се нарича улеснена дифузия; протича с участието на белтъци-носители. Улесненият процес на дифузия не изисква енергия. По този начин се транспортират захари, аминокиселини и азотни основи. Този процес се случва например, когато захарите се абсорбират от чревния лумен от епителните клетки.
Преносът на молекули и йони срещу електрохимичен градиент (активен транспорт) е свързан със значителни енергийни разходи. Градиентите често достигат големи стойности. например концентрационният градиент на водородните йони върху плазмената мембрана на клетките на стомашната лигавица е 106, концентрационният градиент на калциевите йони върху мембраната на саркоплазмения ретикулум е 104, докато потокът от йони срещу градиента е значителен. В резултат на това енергийните разходи за транспортни процеси достигат, например, при хората повече от 1/3 от общата метаболитна енергия. В плазмените мембрани на клетките на различни органи са открити активни транспортни системи за натриеви и калиеви йони - натриева помпа. Тази система изпомпва натрий от клетката и калий в клетката (антипорт) срещу техните електрохимични градиенти. Йонният транспорт се осъществява от основния компонент на натриевата помпа - Na+, K+-зависима АТФ-аза поради хидролиза на АТФ. За всяка хидролизирана ATP молекула се транспортират три натриеви йона и два калиеви йона. Има два вида Са2+-АТФази. Единият от тях осигурява освобождаването на калциеви йони от клетката в междуклетъчната среда, другият осигурява натрупването на калций от клетъчното съдържание във вътреклетъчното депо. И двете системи са в състояние да създадат значителен градиент на калциевите йони. K+, H+-ATPase се намира в лигавицата на стомаха и червата. Той е способен да транспортира Н+ през мембраната на лигавичните везикули по време на хидролиза на АТФ.
Статия: Транспорт на вещества през биологични мембрани
Анион-чувствителна АТФ-аза е открита в микрозоми на лигавицата на стомаха на жаба, която е способна да антипортира бикарбонат и хлорид по време на хидролиза на АТФ.
Описаните механизми на транспортиране на различни вещества през клетъчните мембрани се осъществяват и в случай на транспортирането им през епитела на редица органи (черва, бъбреци, бели дробове), което се осъществява през слой от клетки (монослой в червата и нефроните) , а не през една клетъчна мембрана. Този вид транспорт се нарича трансцелуларен или трансепителен. Характерна особеност на клетките, например чревните епителни клетки и нефронните тубули, е, че техните апикални и базални мембрани се различават по пропускливост, мембранен потенциал и транспортна функция.
Способност за генериране на биоелектрични потенциали и провеждане на възбуждане. Появата на биоелектрични потенциали е свързана със структурните особености на биологичните мембрани и с активността на техните транспортни системи, които създават неравномерно разпределение на йони от двете страни на мембраната (виж. Биоелектрични потенциали, Възбуждане).
Процеси на трансформация и съхранение на енергияпротичат в специализирани биологични мембрани и заемат централно място в енергоснабдяването на живите системи. Двата основни процеса на производство на енергия - фотосинтезата и тъканното дишане - са локализирани в мембраните на вътреклетъчните органели на висшите организми, а при бактериите - в клетъчната (плазмената) мембрана (виж. Тъканно дишане). Фотосинтетичните мембрани преобразуват светлинната енергия в енергията на химичните съединения, съхранявайки я под формата на захари - основният химичен източник на енергия за хетеротрофните организми. По време на дишането енергията на органичните субстрати се освобождава в процеса на пренос на електрони по веригата от редокс транспортери и се използва в процеса на фосфорилиране на ADP от неорганичен фосфат за образуване на ATP. Мембраните, които извършват фосфорилиране, свързано с дишането, се наричат конюгиращи (вътрешни мембрани на митохондриите, клетъчни мембрани на някои аеробни бактерии, мембрани на хроматофори на фотосинтезиращи бактерии).
Метаболитни функциимембраните се определят от два фактора: първо, връзката на голям брой ензими и ензимни системи с мембрани, и второ, способността на мембраните физически да разделят клетката на отделни отделения, разграничавайки метаболитните процеси, протичащи в тях един от друг. Метаболитните системи не остават напълно изолирани. Мембраните, разделящи клетката, съдържат специални системи, които осигуряват селективно навлизане на субстрати, освобождаване на продукти и движение на съединения, които имат регулаторен ефект.
Клетъчна рецепция и междуклетъчни взаимодействия. Тази формула съчетава много обширен и разнообразен набор от важни функции на клетъчните мембрани, които определят взаимодействието на клетката с околната среда и формирането на многоклетъчен организъм като едно цяло. Молекулярно-мембранните аспекти на клетъчното приемане и междуклетъчните взаимодействия се отнасят предимно до имунните реакции, хормоналния контрол на растежа и метаболизма и моделите на ембрионалното развитие.
Нарушения в структурата и функцията на биологичните мембрани. Разнообразието от видове биологични мембрани, тяхната многофункционалност и висока чувствителност към външни условия пораждат изключително разнообразие от структурни и функционални мембранни нарушения, които възникват от много неблагоприятни ефекти и са свързани с огромен брой специфични заболявания на тялото като цяло. . Цялото това разнообразие от нарушения може да бъде условно разделено на транспортни, функционално-метаболитни и структурни. Не е възможно да се характеризира общо последователността на възникване на тези нарушения и във всеки конкретен случай е необходим подробен анализ, за да се изясни основната връзка във веригата на развитие на структурни и функционални мембранни нарушения. Нарушаването на мембранните транспортни функции, по-специално повишената пропускливост на мембраната, е добре известен универсален признак за клетъчно увреждане. Нарушаването на транспортните функции (например при хората) се причинява от повече от 20 така наречени транспортни заболявания, включително бъбречна глюкозурия, цистинурия, нарушена абсорбция на глюкоза, галактоза и витамин В12, наследствена сфероцитоза и др. Сред функционалните и метаболитни нарушения на биологични мембрани, промените в процесите на биосинтеза са централни, както и различни отклонения в енергийното снабдяване на живите системи. В най-общата форма последствието от тези процеси е нарушение на състава и физикохимичните свойства на мембраните, загубата на отделни части от метаболизма и неговото изкривяване, както и намаляване на нивото на жизненоважни енергозависими процеси (активни транспорт на йони, процеси на свързан транспорт, функциониране на контрактилни системи и др.). Увреждането на ултраструктурната организация на биологичните мембрани се изразява в прекомерно образуване на везикули, увеличаване на повърхността на плазмените мембрани поради образуването на мехурчета и процеси, сливане на различни клетъчни мембрани, образуване на микропори и локални структурни дефекти.
Библиография:Биологични мембрани, изд. Д.С. Парсънс, прев. от англ., М., 1978; Болдирев А.А. Въведение в биохимията на мембраните, М., 1986, библиогр.; Конев С.В. и Мажул В.М. Междуклетъчни контакти. Минск, 1977; Кулберг А.Я. Рецептори на клетъчните мембрани, М., 1987, библиогр.; Маленков А.Г. и Чуич Г.А. Междуклетъчни контакти и тъканни реакции, М., 1979; Сим Е . Биохимия на мембраните, прев. от англ., М., 1985, библиогр.; Finean J., Coleman R. и Mitchell R. Мембрани и техните функции в клетката, прев. от англ., М., 1977, библиогр.
внимание! статия " Биологични мембрани“ се предоставя само за информационни цели и не трябва да се използва за самолечение
Транспорт на вещества през плазмената мембрана
Бариерно-транспортната функция на апарата на клетъчната повърхност се осигурява чрез селективен трансфер на йони, молекули и надмолекулни структури в и извън клетката. Транспортът през мембраните осигурява доставка на хранителни вещества и отстраняване на крайните метаболитни продукти от клетката, секреция, създаване на йонни градиенти и трансмембранен потенциал, поддържане на необходимите стойности на pH в клетката и др.
Механизмите на транспортиране на веществата в и извън клетката зависят от химическа природа транспортирано вещество и неговите концентрации от двете страни на клетъчната мембрана, както и от размера транспортирани частици. Малките молекули и йони се транспортират през мембраната чрез пасивен или активен транспорт. Прехвърлянето на макромолекули и големи частици се извършва чрез транспортиране в „мембранна опаковка“, т.е. поради образуването на мехурчета, заобиколени от мембрана.
![]()
Пасивен транспортсе нарича пренасяне на вещества през мембрана по градиента на тяхната концентрация без разход на енергия. Такъв транспорт се осъществява чрез два основни механизма: проста дифузия и улеснена дифузия.
от проста дифузиятранспортират се малки полярни и неполярни молекули, мастни киселини и други нискомолекулни хидрофобни органични вещества. Транспортът на водните молекули през мембраната, осъществяван чрез пасивна дифузия, се нарича осмоза.Пример за проста дифузия е транспортирането на газове през плазмената мембрана на ендотелните клетки на кръвоносните капиляри в околната тъканна течност и обратно.
Хидрофилните молекули и йони, които не могат самостоятелно да преминат през мембраната, се транспортират с помощта на специфични мембранни транспортни протеини. Този транспортен механизъм се нарича улеснена дифузия.
Има два основни класа мембранни транспортни протеини: протеини носителиИ канални протеини.Молекулите на транспортираното вещество, свързващи се с протеин носителпричиняват неговите конформационни промени, което води до прехвърляне на тези молекули през мембраната. Улеснената дифузия се характеризира с висока селективност по отношение на транспортираните вещества.
Канални протеиниобразуват пълни с вода пори, които проникват през липидния двоен слой. Когато тези пори са отворени, неорганични йони или молекули на транспортирани вещества преминават през тях и по този начин се транспортират през мембраната. Йонните канали транспортират приблизително 106 йона в секунда, което е повече от 100 пъти скоростта на транспортиране, извършвано от протеините носители.
Повечето канални протеини имат "порти",които се отварят за кратко и след това се затварят. В зависимост от естеството на канала, портите могат да се отворят в отговор на свързването на сигнални молекули (лиганд-зависими канали), промени в мембранния потенциал (волтаж-зависими канали) или механична стимулация.
Активен транспортсе нарича пренасяне на вещества през мембрана срещу техните градиенти на концентрация. Осъществява се с помощта на протеини-носители и изисква енергия, чийто основен източник е АТФ.
Пример за активен транспорт, който използва енергията на хидролизата на АТФ, за да изпомпва Na+ и K+ йони през клетъчната мембрана, е работата натриево-калиева помпа, осигурявайки създаването на мембранен потенциал върху плазмената мембрана на клетките.
![]()
Помпата се образува от специфични протеинови ензими аденозинтрифосфатази, вградени в биологични мембрани, които катализират разцепването на остатъците от фосфорна киселина от молекулата на АТФ. АТФазите включват: ензимен център, йонен канал и структурни елементи, които предотвратяват обратното изтичане на йони по време на работа на помпата. Работата на натриево-калиевата помпа изисква повече от 1/3 от АТФ, консумиран от клетката.
В зависимост от способността на транспортните протеини да транспортират един или повече видове молекули и йони, пасивният и активният транспорт се делят на унипорт и копорт, или свързан транспорт.
![]()
Унипорт -Това е транспорт, при който протеинът носител функционира само по отношение на молекули или йони от един тип. При копорт, или свързан транспорт, протеинът носител е способен да транспортира два или повече вида молекули или йони едновременно. Тези протеини-носители се наричат съпортьори, или свързани превозвачи.Има два вида копорт: simport и antiport. Кога симпортамолекули или йони се транспортират в една посока и кога антипорт -в противоположни посоки. Например, натриево-калиевата помпа работи на принципа на антипорта, като активно изпомпва Na+ йони от клетките и K+ йони в клетките срещу техните електрохимични градиенти. Пример за симпорт е реабсорбцията на глюкоза и аминокиселини от първичната урина от бъбречните тубулни клетки. В първичната урина концентрацията на Na+ винаги е значително по-висока, отколкото в цитоплазмата на клетките на бъбречните тубули, което се осигурява от работата на натриево-калиевата помпа. Свързването на първичната глюкоза в урината с конюгирания протеин-носител отваря Na+ канала, което се придружава от прехвърлянето на Na+ йони от първичната урина в клетката по техния концентрационен градиент, т.е. чрез пасивен транспорт. Потокът от Na+ йони, от своя страна, причинява промени в конформацията на носещия протеин, което води до транспортиране на глюкоза в същата посока като Na+ йони: от първичната урина в клетката. В този случай, както се вижда, за транспортирането на глюкоза спрегнатият носител използва енергията на Na+ йонния градиент, създаден от работата на натриево-калиевата помпа. Така работата на натриево-калиевата помпа и свързания транспортер, който използва градиент от Na+ йони за транспортиране на глюкоза, прави възможно реабсорбирането на почти цялата глюкоза от първичната урина и включването й в общия метаболизъм на тялото.
Благодарение на селективния транспорт на заредени йони, плазмалемата на почти всички клетки носи положителни заряди от външната си страна и отрицателни заряди от вътрешната си цитоплазмена страна. В резултат на това се създава потенциална разлика между двете страни на мембраната.
Образуването на трансмембранния потенциал се постига главно благодарение на работата на транспортните системи, вградени в плазмалемата: натриево-калиевата помпа и протеиновите канали за K+ йони.
Както беше отбелязано по-горе, по време на работата на натриево-калиевата помпа, за всеки два калиеви йона, абсорбирани от клетката, три натриеви йона се отстраняват от нея. В резултат на това извън клетките се създава излишък от Na+ йони, а вътре в тях се създава излишък от K+ йони. Но още по-значим принос за създаването на трансмембранния потенциал имат калиевите канали, които винаги са отворени в клетките в покой. Поради това K+ йони се движат по градиент на концентрация от клетката в извънклетъчната среда. В резултат на това между двете страни на мембраната възниква потенциална разлика от 20 до 100 mV. Плазмената мембрана на възбудимите клетки (нервни, мускулни, секреторни), заедно с K+ каналите, съдържа множество Na+ канали, които се отварят за кратко време, когато клетката е изложена на химични, електрически или други сигнали. Отварянето на Na+ каналите предизвиква промяна в трансмембранния потенциал (мембранна деполяризация) и специфичен клетъчен отговор на сигнала.
Наричат се транспортни протеини, които генерират потенциални разлики през мембраната електрогенни помпи.Натриево-калиевата помпа служи като основна електрогенна помпа на клетките.
Транспорт в мембранна опаковкахарактеризиращ се с факта, че транспортираните вещества на определени етапи от транспорта се намират вътре в мембранните везикули, т.е. те са заобиколени от мембрана. В зависимост от посоката, в която се транспортират веществата (вътре или извън клетката), транспортът в мембранна опаковка се разделя на ендоцитоза и екзоцитоза.
Ендоцитозае процес на абсорбция от клетка на макромолекули и по-големи частици (вируси, бактерии, клетъчни фрагменти). Ендоцитозата се осъществява чрез фагоцитоза и пиноцитоза.
Фагоцитоза -процесът на активно улавяне и абсорбция от клетка на твърди микрочастици, чийто размер е повече от 1 микрон (бактерии, клетъчни фрагменти и др.). По време на фагоцитозата клетката с помощта на специални рецептори разпознава специфични молекулни групи на фагоцитираната частица.
![]()
![]()
След това в точката на контакт на частицата с клетъчната мембрана се образуват израстъци на плазмалемата - псевдоподия,които обгръщат микрочастицата от всички страни. В резултат на сливането на псевдоподии, такава частица е затворена вътре в везикула, заобиколена от мембрана, която се нарича фагозома.Образуването на фагозоми е енергийно зависим процес и протича с участието на актомиозиновата система. Фагозомата, потапяйки се в цитоплазмата, може да се слее с късна ендозома или лизозома, в резултат на което се усвоява органична микрочастица, абсорбирана от клетката, например бактериална клетка. При хората само няколко клетки са способни на фагоцитоза: например макрофагите на съединителната тъкан и кръвните левкоцити. Тези клетки абсорбират бактерии, както и различни твърди частици, които влизат в тялото, и по този начин го предпазват от патогени и чужди частици.
Пиноцитоза- абсорбция на течност от клетката под формата на истински и колоидни разтвори и суспензии. Този процес като цяло е подобен на фагоцитозата: капка течност се потапя в образуваната вдлъбнатина на клетъчната мембрана, заобиколена от нея и се оказва затворена във везикула с диаметър 0,07-0,02 микрона, потопена в хиалоплазмата на клетката.
Механизмът на пиноцитозата е много сложен. Този процес протича в специализирани зони на апарата на клетъчната повърхност, наречени оградени ями, които заемат около 2% от клетъчната повърхност. Оградени ямиса малки инвагинации на плазмалемата, до които има голямо количество протеин в периферната хиалоплазма клатрин.В областта на оградените ями на повърхността на клетките също има множество рецептори, които могат специфично да разпознават и свързват транспортираните молекули. Когато рецепторите свържат тези молекули, настъпва полимеризация на клатрина и плазмалемата инвагинира. Като резултат, ограден балон,пренасящи транспортирани молекули. Тези мехурчета са получили името си поради факта, че клатринът на тяхната повърхност под електронен микроскоп изглежда като неравна граница.
Транспорт на вещества през биомембрани
След отделяне от плазмалемата, оградените везикули губят клатрин и придобиват способността да се сливат с други везикули. Процесите на полимеризация и деполимеризация на клатрина изискват енергия и се блокират при липса на АТФ.
Пиноцитозата, поради високата концентрация на рецептори в оградените ямки, осигурява селективността и ефективността на транспорта на специфични молекули. Например, концентрацията на молекули на транспортирани вещества в оградени ями е 1000 пъти по-висока от концентрацията им в околната среда. Пиноцитозата е основният метод за транспорт на протеини, липиди и гликопротеини в клетката. Чрез пиноцитозата клетката абсорбира количество течност, равно на нейния обем на ден.
Екзоцитоза- процесът на отстраняване на вещества от клетката. Веществата, които трябва да бъдат отстранени от клетката, първо се затварят в транспортни везикули, чиято външна повърхност обикновено е покрита с протеин клатрин, след което такива везикули се насочват към клетъчната мембрана. Тук мембраната на везикулите се слива с плазмалемата и съдържанието им се излива извън клетката или, като поддържа връзка с плазмалемата, се включва в гликокаликса.
Има два вида екзоцитоза: конститутивна (основна) и регулирана.
Конститутивна екзоцитозапротича непрекъснато във всички клетки на тялото. Той служи като основен механизъм за отстраняване на метаболитни продукти от клетката и постоянно възстановяване на клетъчната мембрана.
Регулирана екзоцитозасе извършва само в специални клетки, които изпълняват секреторна функция. Секретираният секрет се натрупва в секреторни везикули и екзоцитозата настъпва само след като клетката получи съответния химичен или електрически сигнал. Например, β-клетките на островите на Лангерханс в панкреаса освобождават секретите си в кръвта само когато концентрацията на глюкоза в кръвта се повиши.
По време на екзоцитоза секреторните везикули, образувани в цитоплазмата, обикновено се насочват към специализирани области на повърхностния апарат, съдържащи голям брой слети протеини или слети протеини. Когато слетите протеини на плазмената мембрана и секреторната везикула взаимодействат, се образува слята пора, свързваща кухината на везикула с извънклетъчната среда. В този случай се активира актомиозиновата система, в резултат на което съдържанието на везикула се излива от нея извън клетката. По този начин, по време на индуцирана екзоцитоза, енергията е необходима не само за транспортирането на секреторни везикули до плазмалемата, но и за процеса на секреция.
Трансцитоза, или отдих , - Това е транспорт, при който отделните молекули се пренасят през клетката. Този вид транспорт се постига чрез комбинация от ендо- и екзоцитоза. Пример за трансцитоза е транспортирането на вещества през клетките на съдовите стени на човешките капиляри, което може да се случи както в едната, така и в другата посока.
БИОФИЗИКА НА ТРАНСПОРТА НА ВЕЩЕСТВОТО ПРЕЗ МЕМБРАНАТА.
Въпроси за самопроверка
1. Какви обекти включва инфраструктурата на автотранспортния комплекс?
2. Посочете основните компоненти на замърсяването на околната среда от автомобилния транспортен комплекс.
3. Назовете основните причини за образуването на замърсяване на околната среда от автомобилния транспортен комплекс.
4. Назовете източниците, опишете механизмите на образуване и характеризирайте състава на замърсяването на въздуха от промишлени зони и райони на предприятията на автомобилния транспорт.
5. Дайте класификация на отпадъчните води от предприятията на автомобилния транспорт.
6. Назовете и характеризирайте основните замърсители на отпадъчните води от предприятията на автомобилния транспорт.
7. Опишете проблема с промишлените отпадъци от предприятията на автомобилния транспорт.
8. Характеризирайте разпределението на масата на вредните емисии и отпадъците от ATK по техните видове.
9. Анализирайте приноса на инфраструктурните съоръжения на АТК за замърсяването на околната среда.
10. Какви видове стандарти съставляват системата от екологични стандарти. Опишете всеки от тези видове стандарти.
| 1. Бондаренко Е.В. Екологична безопасност на автомобилния транспорт: учебник за университети / E.V. Бондаренко, А.Н. Новиков, А.А. Филипов, О.В. Чекмарева, В.В. Василиева, М.В. Коротков // Орел: Орловски държавен технически университет, 2010. – 254 с. 2. Бондаренко Е.В. Екология на автомобилния транспорт: [Текст]: учеб. помощ / E.V. Бондаренко, Г.П. Дворников Оренбург: РИК ГОУ ОСУ, 2004. – 113 с. 3. Каганов И.Л. Наръчник по санитария и хигиена в автотранспортните предприятия. [Текст] / I.L. Каганов, В. Д. Морошек Мн.: Беларус, 1991. – 287 с. 4. Картошкин А.П. Концепцията за събиране и преработка на използвани смазочни масла / A.P. Картошкин // Химия и технология на горива и масла, 2003. - № 4. – С. 3 – 5. 5. Луканин В.Н. Промишлена и транспортна екология [Текст] / V.N. Луканин, Ю.В. Трофименко М.: Висше. училище, 2001. - 273 с. 6. Руска енциклопедия на автомобилния транспорт. Техническа експлоатация, поддръжка и ремонт на автомобили. – Т.3. – М.: RBOOIP “Просвещение”, 2001. – 456 с. | |
Клетката е отворена система, която непрекъснато обменя материя и енергия с околната среда. Преносът на вещества през биологичните мембрани е необходимо условие за живота. С преноса на вещества през мембраните са свързани клетъчните метаболитни процеси, биоенергийните процеси, образуването на биопотенциали, генерирането на нервен импулс и др.. Нарушаването на транспорта на вещества през биомембраните води до различни патологии. Лечението често включва проникване на лекарства през клетъчните мембрани. Клетъчната мембрана е селективна бариера за различни вещества, намиращи се вътре и извън клетката. Има два вида мембранен транспорт: пасивен и активен транспорт.
всичко видове пасивен транспортна принципа на дифузията. Дифузията е резултат от хаотични независими движения на много частици. Дифузията постепенно намалява концентрационния градиент до достигане на състояние на равновесие. В този случай във всяка точка ще се установи еднаква концентрация и дифузията в двете посоки ще се случи еднакво.Дифузията е пасивен транспорт, тъй като не изисква външна енергия. Има няколко вида дифузия в плазмената мембрана:
1 ) Свободна дифузия.
123456Следващ ⇒
Прочетете също:
Видео: Транспорт в клетки Дифузия и осмоза, част - 1 Транспорт в клетки: Дифузия и осмоза, част - 1
дифузияпрез клетъчната мембрана се разделят на два подтипа: проста дифузия и улеснена дифузия. Простата дифузия означава, че кинетичното движение на молекули или йони се осъществява през мембранен отвор или междумолекулни пространства без никакво взаимодействие с мембранни протеини-носители. Скоростта на дифузия се определя от количеството вещество, скоростта на кинетичното движение и броя и размера на отворите в мембраната, през които могат да се движат молекули или йони.
Видео: Транспорт на вещества в тялото
Улеснена дифузияизисква взаимодействие с протеин-носител, който улеснява транспорта на молекули или йони чрез химично свързване с тях и в тази форма преминава през мембраната.
Проста дифузияможе да се появи през клетъчната мембрана по два начина: (1) през междумолекулните пространства на липидния двоен слой, ако дифузиращото вещество е мастноразтворимо; (2) през пълни с вода канали, които проникват в някои големи транспортни протеини, както е показано на фиг.
Пренос на вещества през мембраната. Активен и пасивен транспорт на вещества през мембраната
Дифузия на мастноразтворими веществапрез липидния двоен слой. Един от най-важните фактори, определящи скоростта на дифузия на дадено вещество през липиден бислой, е неговата разтворимост в липиди. Например, кислородът, азотът, въглеродният диоксид и алкохолите имат по-висока разтворимост в липидите, така че те могат директно да се разтварят в липидния двоен слой и да дифундират през клетъчната мембрана по същия начин, по който водоразтворимите вещества дифундират във водни разтвори. Очевидно е, че степента на дифузия на всяко от тези вещества е правопропорционална на тяхната разтворимост в липиди. По този начин могат да се транспортират много големи количества кислород. По този начин кислородът може да бъде доставен в клетките почти толкова бързо, колкото ако клетъчната мембрана не съществува.
Дифузия на вода и други неразтворими в мазнини веществамолекули през протеинови канали. Въпреки факта, че водата изобщо не се разтваря в липидите на мембраната, тя лесно преминава през каналите в протеиновите молекули, които проникват през мембраната. Скоростта, с която водните молекули могат да се движат през повечето клетъчни мембрани, е невероятна. Например, общото количество вода, което дифундира във всяка посока през мембраната на червени кръвни клетки за секунда, е около 100 пъти обема на самата клетка.
Чрез представените канали протеинови пори, други неразтворими в липиди молекули могат да преминат, ако са водоразтворими и достатъчно малки. Въпреки това, увеличаването на размера на такива молекули бързо намалява тяхната проникваща способност. Например способността на уреята да проникне през мембраната е приблизително 1000 пъти по-малка от тази на водата, въпреки че диаметърът на молекулата на уреята е само 20% по-голям от диаметъра на водната молекула. Въпреки това, като се има предвид невероятната скорост на преминаване на водата, пропускливостта на уреята осигурява бързия й транспорт през мембраната в рамките на няколко минути.
Дифузия през протеинови канали
Компютър триизмерен реконструкция на протеинови каналидемонстрира наличието на тубулни структури, които проникват през мембраната през и през - от извънклетъчната към вътреклетъчната течност. Следователно веществата могат да се движат през тези канали чрез проста дифузия от едната страна на мембраната към другата. Протеиновите канали се отличават с две важни характеристики: (1) те често са селективно пропускливи за определени вещества; (2) много канали могат да бъдат отворени или затворени от порти.
Видео: Мембранни потенциали - Част 1
Изборен пропускливост на протеинови канали. Много протеинови канали са силно селективни за транспортирането на един или повече специфични йони или молекули. Това се дължи на собствените характеристики на канала (диаметър и форма), както и на естеството на електрическите заряди и химическите връзки на повърхностите, които го покриват. Например, един от най-важните протеинови канали - така нареченият натриев канал - има диаметър от 0,3 до 0,5 nm, но по-важното е, че вътрешните повърхности на този канал са силно отрицателно заредени. Тези отрицателни заряди могат да привлекат малки, дехидратирани натриеви йони в каналите, като по същество издърпват тези йони далеч от водните молекули, които ги заобикалят. Веднъж попаднали в канала, натриевите йони дифундират във всяка посока според обичайните правила за дифузия. В това отношение натриевият канал е специфично селективен за провеждането на натриеви йони.
Тези канали са малко по-малки от натриевите канали канали, диаметърът им е само около 0,3 nm, но те не са отрицателно заредени и имат различни химични връзки. Следователно няма изразена сила, която да издърпва йоните в канала, и калиевите йони не се освобождават от тяхната водна обвивка. Хидратираната форма на калиевия йон е много по-малка по размер от хидратираната форма на натриевия йон, тъй като натриевият йон привлича много повече водни молекули от калиевия йон. Следователно по-малките хидратирани калиеви йони могат лесно да преминат през този тесен канал, докато по-големият хидратиран натриев йон се „отстранява“, позволявайки селективна пропускливост за конкретен йон.
Източник: http://meduniver.com
Внимание, само ДНЕС!
Транспорт на вещества: механизми на проникване на вещества в клетката
Пасивен транспорт
Движението на вещество (йони или малки молекули) по градиент на концентрация. Осъществява се без разход на енергия чрез проста дифузия, осмоза или улеснена дифузия с помощта на протеини-носители.
Активен транспорт
Транспорт на вещества (йони или малки молекули) с помощта на протеини носители срещу концентрационен градиент. Извършва се с цената на ATP.
Ендоцитоза
Абсорбция на вещества (големи частици или макромолекули) чрез обграждането им с израстъци на цитоплазмената мембрана с образуване на заобиколени от мембрана везикули.
Екзоцитоза
Освобождаването на вещества (големи частици или макромолекули) от клетката чрез обграждането им с израстъци на цитоплазмената мембрана с образуването на заобиколени от мембрана везикули.
Фагоцитоза и обратна фагоцитоза
Абсорбция и освобождаване на твърди и големи частици. Характерен за животински и човешки клетки.
Пиноцитоза и обратна пиноцитоза
Абсорбция и освобождаване на течни и разтворени частици. Характерен за растителни и животински клетки.
Кириленко А. А. Биология.
ТРАНСПОРТ НА ВЕЩЕСТВАТА ПРЕЗ МЕМБРАНАТА
Единен държавен изпит. Секция "Молекулярна биология". Теория, тренировъчни задачи. 2017 г.
химическа природа транспортирано вещество и неговите концентрации от размери

Пасивен транспорт
от проста дифузия осмоза.
улеснена дифузия.
протеини носителиИ канални протеини. протеин носител
Канални протеини
"порти",които се отварят за кратко и след това се затварят.
В зависимост от естеството на канала, портата може да се отвори в отговор на свързването на сигнални молекули (лиганд-зависими канали), промени в мембранния потенциал (волтаж-зависими канали) или механична стимулация.
Активен транспорт
натриево-калиева помпа

Помпата се образува от специфични ензимни протеини аденозин трифосфатаза, вградени в биологични мембрани, които катализират разцепването на остатъците от фосфорна киселина от молекулата на АТФ.
АТФазите включват: ензимен център, йонен канал и структурни елементи, които предотвратяват обратното изтичане на йони по време на работа на помпата. Повече от 1/3 от АТФ, консумиран от клетката, се изразходва за работата на натриево-калиевата помпа.

Унипорт - съпортьори, или свързани превозвачи. симпорта антипорт -в противоположни посоки. Например, натриево-калиевата помпа работи на принципа на антипорта, като активно изпомпва Na + йони от клетките и K + йони в клетките срещу техните електрохимични градиенти. Пример за симпорт е реабсорбцията на глюкоза и аминокиселини от първичната урина от бъбречните тубулни клетки. В първичната урина концентрацията на Na + винаги е значително по-висока, отколкото в цитоплазмата на бъбречните тубулни клетки, което се осигурява от работата на натриево-калиевата помпа. Свързването на първичната глюкоза в урината с конюгирания протеин-носител отваря Na + канала, който се придружава от прехвърлянето на Na + йони от първичната урина в клетката по техния концентрационен градиент, т.е. чрез пасивен транспорт. Потокът от Na + йони, от своя страна, причинява промени в конформацията на протеина носител, което води до транспортиране на глюкоза в същата посока като Na + йони: от първичната урина в клетката.
В този случай за транспортирането на глюкоза, както може да се види, конюгатният транспортер използва енергията на Na + йонния градиент, създаден от работата на натриево-калиевата помпа. По този начин работата на натриево-калиевата помпа и свързания транспортер, който използва градиент от Na + йони за транспортиране на глюкоза, прави възможно реабсорбирането на почти цялата глюкоза от първичната урина и включването й в общия метаболизъм на тялото.
Както беше отбелязано по-горе, по време на работата на натриево-калиевата помпа, за всеки два калиеви йона, абсорбирани от клетката, три натриеви йона се отстраняват от нея. В резултат на това извън клетките се създава излишък от Na + йони, а вътре - излишък от K + йони. Но още по-значим принос за създаването на трансмембранния потенциал имат калиевите канали, които винаги са отворени в клетките в покой. Поради това K+ йони излизат от клетката по концентрационен градиент в извънклетъчната среда. В резултат на това между двете страни на мембраната възниква потенциална разлика от 20 до 100 mV. Плазмената мембрана на възбудимите клетки (нервни, мускулни, секреторни), заедно с K + каналите, съдържа множество Na + канали, които се отварят за кратко време, когато върху клетката действат химически, електрически или други сигнали. Отварянето на Na+ каналите предизвиква промяна в трансмембранния потенциал (мембранна деполяризация) и специфичен клетъчен отговор на сигнала.
електрогенни помпи.
характеризиращ се с факта, че транспортираните вещества на определени етапи от транспорта се намират вътре в мембранните везикули, т.е. те са заобиколени от мембрана.
22. Транспорт на вещества през мембраната. Активен и пасивен транспорт
В зависимост от посоката, в която се транспортират веществата (вътре или извън клетката), транспортът в мембранна опаковка се разделя на ендоцитоза и екзоцитоза.
Ендоцитоза
Фагоцитоза -


псевдоподия, фагозома.
Пиноцитоза
Оградени ями клатрин. ограден балон,
Екзоцитоза
Конститутивна екзоцитоза
Регулирана екзоцитоза
По време на екзоцитоза секреторните везикули, образувани в цитоплазмата, обикновено се насочват към специализирани области на повърхностния апарат, съдържащи голям брой слети протеини или слети протеини. Когато слетите протеини на плазмената мембрана и секреторната везикула взаимодействат, се образува слята пора, свързваща кухината на везикула с извънклетъчната среда. В този случай се активира актомиозиновата система, в резултат на което съдържанието на везикула се излива от нея извън клетката. По този начин, по време на индуцируема екзоцитоза, енергията е необходима не само за транспортирането на секреторни везикули до плазмалемата, но и за процеса на секреция.
Трансцитоза, или отдих , -
Методи за транспортиране на вещества през мембрана.
Повечето жизненоважни процеси, като абсорбция, екскреция, провеждане на нервни импулси, мускулна контракция, синтез на АТФ, поддържане на постоянен йонен състав и водно съдържание са свързани с преноса на вещества през мембраните. Този процес в биологичните системи се нарича транспорт . Обменът на вещества между клетката и околната среда се извършва постоянно. Механизмите на транспортиране на вещества в и извън клетката зависят от размера на транспортираните частици. Малките молекули и йони се транспортират от клетката директно през мембраната под формата на пасивен и активен транспорт.
Пасивен транспортизвършва се без разход на енергия, по градиент на концентрация чрез проста дифузия, филтрация, осмоза или улеснена дифузия.
дифузия – проникване на вещества през мембраната по концентрационен градиент (от област, където концентрацията им е по-висока, към област, където концентрацията им е по-ниска); този процес протича без консумация на енергия поради хаотичното движение на молекулите. Дифузният транспорт на вещества (вода, йони) се осъществява с участието на интегрални мембранни протеини, които имат молекулни пори (канали, през които преминават разтворени молекули и йони), или с участието на липидната фаза (за мастноразтворимите вещества) . С помощта на дифузия в клетката проникват разтворени молекули на кислород и въглероден диоксид, както и отрови и лекарства.

Видове транспорт през мембраната: 1 – проста дифузия; 2 – дифузия през мембранни канали; 3 – улеснена дифузия с помощта на протеини-носители; 4 – активен транспорт.
Улеснена дифузия. Транспортът на веществата през липидния двоен слой чрез проста дифузия се извършва с ниска скорост, особено в случай на заредени частици, и е почти неконтролиран. Следователно в процеса на еволюция за някои вещества се появяват специфични мембранни канали и мембранни транспортери, които спомагат за увеличаване на скоростта на пренос и освен това извършват селективентранспорт.
Пасивен транспорт на вещества с помощта на носители се нарича улеснена дифузия. В мембраната са вградени специални протеини-носители (пермеаза). Пермеазите селективно се свързват с един или друг йон или молекула и ги пренасят през мембраната. В този случай частиците се движат по-бързо, отколкото при конвенционалната дифузия.
Осмоза – навлизане на вода в клетките от хипотоничен разтвор.
Филтриране — изтичане на вещества от порите към по-ниски стойности на налягането. Пример за филтриране в тялото е прехвърлянето на вода през стените на кръвоносните съдове, изстисквайки кръвната плазма в бъбречните тубули.

Ориз. Движение на катиони по електрохимичен градиент.
Активен транспорт. Ако в клетките съществуваше само пасивен транспорт, тогава концентрациите, наляганията и други стойности извън и вътре в клетката биха били равни. Следователно има друг механизъм, който работи в посока срещу електрохимичния градиент и се получава с разхода на енергия от клетката. Преносът на молекули и йони срещу електрохимичния градиент, осъществяван от клетката благодарение на енергията на метаболитните процеси, се нарича активен транспорт.Той е присъщ само на биологичните мембрани. Активното пренасяне на вещество през мембраната възниква поради свободната енергия, освободена по време на химични реакции вътре в клетката. Активният транспорт в тялото създава градиенти на концентрации, електрически потенциали, налягания, т.е. поддържа живота в тялото.
Активният транспорт се състои от движение на вещества срещу градиент на концентрация с помощта на транспортни протеини (порини, АТФази и др.), образуващи диафрагмени помпи, с разхода на енергия от АТФ (калиево-натриева помпа, регулиране на концентрацията на калциеви и магнезиеви йони в клетките, доставка на монозахариди, нуклеотиди, аминокиселини). Изследвани са 3 основни активни транспортни системи, които осигуряват преноса на Na, K, Ca, H йони през мембраната.
Механизъм. K + и Na + йони са неравномерно разпределени от различните страни на мембраната: концентрацията на Na + отвън > K + йони, а вътре в клетката K + > Na +. Тези йони дифундират през мембраната по посока на електрохимичния градиент, което води до неговото изравняване. Na-K помпите са част от цитоплазмените мембрани и работят благодарение на енергията на хидролиза на ATP молекули с образуването на ADP молекули и неорганичен фосфат F n: ATP=ADP+P n.Помпата работи обратимо: градиентите на концентрация на йони насърчават синтеза на ATP молекули от ADP и Ph n молекули: ADP + Ph n = ATP.
Na + /K + помпата е трансмембранен протеин, способен на конформационни промени, в резултат на което може да прикрепи както "K +", така и "Na +".
Мембранен транспорт
В един цикъл на работа помпата премахва три „Na +“ от клетката и въвежда два „K +“ поради енергията на молекулата на АТФ. Почти една трета от цялата енергия, необходима за функционирането на клетките, се изразходва за работата на натриево-калиевата помпа.


Не само отделни молекули, но и твърди вещества ( фагоцитоза), решения ( пиноцитоза). Фагоцитоза – улавяне и абсорбиране на големи частици(клетки, клетъчни части, макромолекули) и пиноцитоза – улавяне и абсорбиране на течен материал(разтвор, колоиден разтвор, суспензия). Получените пиноцитозни вакуоли варират по размер от 0,01 до 1-2 µm. След това вакуолата се потапя в цитоплазмата и се отделя. В този случай стената на пиноцитозната вакуола напълно запазва структурата на плазмената мембрана, която я е породила.

Ако дадено вещество се транспортира в клетката, тогава този вид транспорт се нарича ендоцитоза (прехвърляне в клетката чрез директно пино или фагоцитоза), ако е навън, тогава – екзоцитоза (прехвърляне от клетката чрез обратно пино или фагоцитоза). В първия случай от външната страна на мембраната се образува инвагинация, която постепенно се превръща във везикула. Везикулата се отделя от мембраната вътре в клетката. Такава везикула съдържа транспортираното вещество, заобиколено от билипидна мембрана (везикула). Впоследствие везикулата се слива с някаква клетъчна органела и освобождава съдържанието си в нея. В случай на екзоцитоза процесът протича в обратен ред: везикулата се приближава към мембраната от вътрешната страна на клетката, слива се с нея и освобождава съдържанието си в междуклетъчното пространство.
Пиноцитозата и фагоцитозата са фундаментално сходни процеси, в които могат да се разграничат четири фази: навлизане на вещества чрез пиноцитоза или фагоцитоза, тяхното разграждане под действието на ензими, секретирани от лизозомите, прехвърляне на продуктите на разпадане в цитоплазмата (поради промени в пропускливостта на вакуолните мембрани) и освобождаването на метаболитни продукти навън. Много протозои и някои левкоцити са способни на фагоцитоза. Пиноцитоза се наблюдава в чревните епителни клетки и в ендотела на кръвоносните капиляри.
Предишен12345678Следващ
ВИЖ ПОВЕЧЕ:
Транспорт на вещества през плазмената мембрана
Бариерната транспортна функция на апарата на клетъчната повърхност се осигурява чрез селективен трансфер на йони, молекули и супрамолекулни структури в и извън клетката. Транспортът през мембраните осигурява доставка на хранителни вещества и отстраняване на крайните метаболитни продукти от клетката, секреция, създаване на йонни градиенти и трансмембранен потенциал, поддържане на необходимите стойности на pH в клетката и др.
Механизмите на транспортиране на веществата в и извън клетката зависят от химическа природа транспортирано вещество и неговите концентрации от двете страни на клетъчната мембрана, както и от размери транспортирани частици. Малките молекули и йони се транспортират през мембраната чрез пасивен или активен транспорт. Прехвърлянето на макромолекули и големи частици се извършва чрез транспортиране в „мембранна опаковка“, т.е. поради образуването на везикули, заобиколени от мембрана.
Пасивен транспортсе нарича пренасяне на вещества през мембрана по техния концентрационен градиент без разход на енергия. Такъв транспорт се осъществява чрез два основни механизма: проста дифузия и улеснена дифузия.
от проста дифузиятранспортират се малки полярни и неполярни молекули, мастни киселини и други нискомолекулни хидрофобни органични вещества. Преносът на водни молекули през мембрана, осъществяван чрез пасивна дифузия, се нарича осмоза.Пример за проста дифузия е транспортирането на газове през плазмената мембрана на ендотелните клетки на кръвоносните капиляри в околната тъканна течност и обратно.
Хидрофилните молекули и йони, които не могат самостоятелно да преминат през мембраната, се транспортират с помощта на специфични мембранни транспортни протеини. Този транспортен механизъм се нарича улеснена дифузия.
Има два основни класа мембранни транспортни протеини: протеини носителиИ канални протеини.Молекулите на транспортираното вещество, свързващи се с протеин носителпричиняват неговите конформационни промени, което води до прехвърляне на тези молекули през мембраната. Улеснената дифузия е силно селективна по отношение на транспортираните вещества.
Канални протеиниобразуват пълни с вода пори, които проникват през липидния двоен слой. Когато тези пори са отворени, неорганичните йони или транспортните молекули преминават през тях и по този начин се транспортират през мембраната. Йонните канали транспортират приблизително 10 6 йона в секунда, което е повече от 100 пъти скоростта на транспортиране, извършвано от протеините носители.
Повечето канални протеини имат "порти",които се отварят за кратко и след това се затварят. В зависимост от естеството на канала, портата може да се отвори в отговор на свързването на сигнални молекули (лиганд-зависими канали), промени в мембранния потенциал (волтаж-зависими канали) или механична стимулация.
Активен транспортсе нарича транспорт на вещества през мембрана срещу градиентите на тяхната концентрация. Осъществява се с помощта на протеини-носители и изисква енергия, чийто основен източник е АТФ.
Пример за активен транспорт, който използва енергията на хидролизата на АТФ, за да изпомпва Na + и K + йони през клетъчната мембрана, е работата натриево-калиева помпа, осигурявайки създаването на мембранен потенциал върху плазмената мембрана на клетките.
Помпата се образува от специфични ензимни протеини аденозин трифосфатаза, вградени в биологични мембрани, които катализират разцепването на остатъците от фосфорна киселина от молекулата на АТФ. АТФазите включват: ензимен център, йонен канал и структурни елементи, които предотвратяват обратното изтичане на йони по време на работа на помпата. Повече от 1/3 от АТФ, консумиран от клетката, се изразходва за работата на натриево-калиевата помпа.
В зависимост от способността на транспортните протеини да транспортират един или повече видове молекули и йони, пасивният и активният транспорт се делят на унипорт и копорт, или свързан транспорт.
Унипорт -Това е транспорт, при който протеинът носител функционира само по отношение на молекули или йони от един тип. При копорт, или свързан транспорт, протеинът носител е способен да транспортира два или повече вида молекули или йони едновременно. Тези протеини-носители се наричат съпортьори, или свързани превозвачи.Има два вида копорт: simport и antiport. Кога симпортамолекули или йони се транспортират в една посока и кога антипорт -в противоположни посоки. Например, натриево-калиевата помпа работи на принципа на антипорта, като активно изпомпва Na + йони от клетките и K + йони в клетките срещу техните електрохимични градиенти.
Пример за симпорт е реабсорбцията на глюкоза и аминокиселини от първичната урина от бъбречните тубулни клетки. В първичната урина концентрацията на Na + винаги е значително по-висока, отколкото в цитоплазмата на бъбречните тубулни клетки, което се осигурява от работата на натриево-калиевата помпа. Свързването на първичната глюкоза в урината с конюгирания протеин-носител отваря Na + канала, който се придружава от прехвърлянето на Na + йони от първичната урина в клетката по техния концентрационен градиент, т.е. чрез пасивен транспорт. Потокът от Na + йони, от своя страна, причинява промени в конформацията на протеина носител, което води до транспортиране на глюкоза в същата посока като Na + йони: от първичната урина в клетката. В този случай за транспортирането на глюкоза, както може да се види, конюгатният транспортер използва енергията на Na + йонния градиент, създаден от работата на натриево-калиевата помпа. По този начин работата на натриево-калиевата помпа и свързания транспортер, който използва градиент от Na + йони за транспортиране на глюкоза, прави възможно реабсорбирането на почти цялата глюкоза от първичната урина и включването й в общия метаболизъм на тялото.
Благодарение на селективния транспорт на заредени йони, плазмалемата на почти всички клетки носи положителни заряди от външната си страна и отрицателни заряди от вътрешната си цитоплазмена страна. В резултат на това се създава потенциална разлика между двете страни на мембраната.
Образуването на трансмембранния потенциал се постига главно благодарение на работата на транспортните системи, вградени в плазмалемата: натриево-калиевата помпа и протеиновите канали за K + йони.
Както беше отбелязано по-горе, по време на работата на натриево-калиевата помпа, за всеки два калиеви йона, абсорбирани от клетката, три натриеви йона се отстраняват от нея. В резултат на това извън клетките се създава излишък от Na + йони, а вътре - излишък от K + йони. Но още по-значим принос за създаването на трансмембранния потенциал имат калиевите канали, които винаги са отворени в клетките в покой. Поради това K+ йони излизат от клетката по концентрационен градиент в извънклетъчната среда. В резултат на това между двете страни на мембраната възниква потенциална разлика от 20 до 100 mV. Плазмената мембрана на възбудимите клетки (нервни, мускулни, секреторни), заедно с K + каналите, съдържа множество Na + канали, които се отварят за кратко време, когато върху клетката действат химически, електрически или други сигнали.
Отварянето на Na+ каналите предизвиква промяна в трансмембранния потенциал (мембранна деполяризация) и специфичен клетъчен отговор на сигнала.
Наричат се транспортни протеини, които генерират потенциални разлики през мембраната електрогенни помпи.Натриево-калиевата помпа служи като основна електрогенна помпа на клетките.
Транспорт в мембранна опаковкахарактеризиращ се с факта, че транспортираните вещества на определени етапи от транспорта се намират вътре в мембранните везикули, т.е. те са заобиколени от мембрана. В зависимост от посоката, в която се транспортират веществата (вътре или извън клетката), транспортът в мембранна опаковка се разделя на ендоцитоза и екзоцитоза.
Ендоцитозае процес на абсорбция от клетка на макромолекули и по-големи частици (вируси, бактерии, клетъчни фрагменти). Ендоцитозата се осъществява чрез фагоцитоза и пиноцитоза.
Фагоцитоза -процесът на активно улавяне и абсорбция от клетка на твърди микрочастици, чийто размер е повече от 1 микрон (бактерии, клетъчни фрагменти и др.). По време на фагоцитозата клетката с помощта на специални рецептори разпознава специфични молекулни групи на фагоцитираната частица.
След това в точката на контакт на частицата с клетъчната мембрана се образуват израстъци на плазмалемата - псевдоподия,които обгръщат микрочастицата от всички страни. В резултат на сливането на псевдоподии, такава частица е затворена вътре в везикула, заобиколена от мембрана, която се нарича фагозома.Образуването на фагозоми е енергийно зависим процес и протича с участието на актомиозиновата система. Фагозомата, потапяйки се в цитоплазмата, може да се слее с късна ендозома или лизозома, в резултат на което органичната микрочастица, абсорбирана от клетката, например бактериална клетка, се усвоява. При хората само няколко клетки са способни на фагоцитоза: например макрофагите на съединителната тъкан и кръвните левкоцити. Тези клетки абсорбират бактерии, както и различни прахови частици, които влизат в тялото, като по този начин го предпазват от патогени и чужди частици.
Пиноцитоза- абсорбция на течност от клетката под формата на истински и колоидни разтвори и суспензии. Този процес като цяло е подобен на фагоцитозата: капка течност се потапя в образуваната вдлъбнатина на клетъчната мембрана, заобиколена от нея и се оказва затворена във везикула с диаметър 0,07-0,02 микрона, потопена в хиалоплазмата на клетката.
Механизмът на пиноцитозата е много сложен. Този процес протича в специализирани зони на повърхностния апарат на клетката, наречени оградени ями, които заемат около 2% от клетъчната повърхност. Оградени ямиса малки инвагинации на плазмалемата, до които има голямо количество протеин в периферната хиалоплазма клатрин.В областта на оградените ями на повърхността на клетките също има множество рецептори, които могат специфично да разпознават и свързват транспортираните молекули. Когато рецепторите свържат тези молекули, настъпва полимеризация на клатрина и плазмалемата инвагинира. Като резултат, ограден балон,пренасящи транспортируеми молекули. Тези мехурчета са получили името си поради факта, че клатринът на тяхната повърхност изглежда като неравен ръб под електронен микроскоп. След отделяне от плазмалемата, оградените везикули губят клатрин и придобиват способността да се сливат с други везикули. Процесите на полимеризация и деполимеризация на клатрина изискват енергия и се блокират при липса на АТФ.
Пиноцитозата, поради високата концентрация на рецептори в оградените ямки, осигурява селективността и ефективността на транспорта на специфични молекули. Например, концентрацията на молекулите на транспортираните вещества в оградените ями е 1000 пъти по-висока от концентрацията им в околната среда. Пиноцитозата е основният метод за транспорт на протеини, липиди и гликопротеини в клетката. Чрез пиноцитозата клетката абсорбира количество течност, равно на нейния обем на ден.
Екзоцитоза- процесът на отстраняване на вещества от клетката. Веществата, които трябва да бъдат отстранени от клетката, първо се затварят в транспортни везикули, чиято външна повърхност обикновено е покрита с протеин клатрин, след което такива везикули се насочват към клетъчната мембрана. Тук мембраната на везикулите се слива с плазмалемата и съдържанието им се излива извън клетката или, като поддържа контакт с плазмалемата, се включва в гликокаликса.
Има два вида екзоцитоза: конститутивна (основна) и регулирана.
Конститутивна екзоцитозапротича непрекъснато във всички клетки на тялото. Той служи като основен механизъм за отстраняване на метаболитни продукти от клетката и постоянно възстановяване на клетъчната мембрана.
Регулирана екзоцитозаизвършва се само в специални клетки, които изпълняват секреторна функция. Секретираният секрет се натрупва в секреторни везикули и екзоцитозата настъпва само след като клетката получи съответния химичен или електрически сигнал. Например, β-клетките на Лангерхансовите острови на панкреаса освобождават своя секрет в кръвта само когато концентрацията на глюкоза в кръвта се повиши.
По време на екзоцитоза секреторните везикули, образувани в цитоплазмата, обикновено се насочват към специализирани области на повърхностния апарат, съдържащи голям брой слети протеини или слети протеини. Когато слетите протеини на плазмената мембрана и секреторната везикула взаимодействат, се образува слята пора, свързваща кухината на везикула с извънклетъчната среда.
В този случай се активира актомиозиновата система, в резултат на което съдържанието на везикула се излива от нея извън клетката. По този начин, по време на индуцируема екзоцитоза, енергията е необходима не само за транспортирането на секреторни везикули до плазмалемата, но и за процеса на секреция.
Трансцитоза, или отдих , - Това е транспорт, при който отделните молекули се пренасят през клетката. Този вид транспорт се постига чрез комбинация от ендо- и екзоцитоза. Пример за трансцитоза е транспортирането на вещества през клетките на съдовите стени на човешките капиляри, което може да се случи както в едната, така и в другата посока.
Състои се от способността му да предава различни вещества в и извън клетката. Това е от голямо значение за саморегулацията и поддържането на постоянен клетъчен състав. Тази функция на клетъчната мембрана се осъществява благодарение на селективна пропускливост, тоест способността да пропуска някои вещества, а не други.
Транспорт през липидния бислой (проста дифузия) и транспорт с участието на мембранни протеини
Неполярните молекули с ниско молекулно тегло (кислород, азот, бензен) преминават най-лесно през липидния бислой. Малки полярни молекули като въглероден диоксид, азотен оксид, вода и урея проникват доста бързо през липидния двоен слой. Етанолът и глицеролът, както и стероидите и хормоните на щитовидната жлеза преминават през липидния двоен слой със забележима скорост. За по-големите полярни молекули (глюкоза, аминокиселини), както и за йоните, липидният бислой е практически непропусклив, тъй като вътрешността му е хидрофобна. Така за вода коефициентът на пропускливост (cm/s) е около 10−2, за глицерол - 10−5, за глюкоза - 10−7, а за едновалентни йони - по-малко от 10−10.
Преносът на големи полярни молекули и йони се осъществява благодарение на канални протеини или протеини-носители. Така в клетъчните мембрани има канали за натриеви, калиеви и хлорни йони, в мембраните на много клетки има водни канали аквапорини, както и протеини-носители за глюкоза, различни групи аминокиселини и много йони.
Активен и пасивен транспорт
Симпорт, антипорт и унипорт
Мембранният транспорт на вещества също се различава по посоката на тяхното движение и количеството вещества, пренасяни от даден носител:
- 1) Унипорт- транспорт на едно вещество в една посока в зависимост от градиента
- 2) Симпорт- транспорт на две вещества в една посока чрез един превозвач.
- 3) Антипорт- движение на две вещества в различни посоки чрез един транспортьор.
Унипортосъществява, например, зависим от напрежението натриев канал, през който натриевите йони се движат в клетката по време на генерирането на потенциал за действие.
Симпортизвършва глюкозен транспортер, разположен от външната (с лице към чревния лумен) страна на чревните епителни клетки. Този протеин едновременно улавя молекула глюкоза и натриев йон и, променяйки конформацията, пренася и двете вещества в клетката. Това използва енергията на електрохимичния градиент, който от своя страна се създава поради хидролизата на АТФ от натриево-калиевата АТФаза.
Антипортизвършва се, например, от натриево-калиева АТФ-аза (или натрий-зависима АТФ-аза). Той транспортира калиеви йони в клетката. а от клетката – натриеви йони.
Работата на натриево-калиевата АТФаза като пример за антипорт и активен транспорт
Първоначално този транспортер свързва три йона към вътрешната страна на мембраната. Тези йони променят конформацията на активния център на ATPase. След такова активиране АТФ-азата е в състояние да хидролизира една АТФ молекула и фосфатният йон се фиксира върху повърхността на транспортера от вътрешната страна на мембраната.
Освободената енергия се изразходва за промяна на конформацията на АТФ-аза, след което три йона N a + (\displaystyle Na^(+))и йонът (фосфатът) завършва от външната страна на мембраната. Ето йоните N a + (\displaystyle Na^(+))се разделят и P O 4 3 − (\displaystyle PO_(4)^(3-))се заменя с два йона. След това конформацията на носителя се променя до първоначалната, а йоните K + (\displaystyle K^(+))се появяват от вътрешната страна на мембраната. Ето йоните K + (\displaystyle K^(+))се отделят и носачът е готов за работа отново.
По-накратко, действията на ATPase могат да бъдат описани, както следва:
В резултат на това се създава висока концентрация на йони в извънклетъчната среда N a + (\displaystyle Na^(+)), а вътре в клетката има висока концентрация K + (\displaystyle K^(+)). работа N a + (\displaystyle Na^(+)), K + (\displaystyle K^(+))- ATPase създава не само разлика в концентрацията, но и разлика в заряда (работи като електрогенна помпа). От външната страна на мембраната се създава положителен заряд, а отвътре - отрицателен.
Активният транспорт на вещества протича срещу общия (генерализиран) градиент. Това означава, че прехвърлянето на вещество става от места с по-ниска стойност на електрохимичния потенциал към места с по-висока стойност.
Активният транспорт не може да възникне спонтанно, а само във връзка с процеса на хидролиза на аденозинтрифосфорната киселина (АТФ), т.е. поради изразходването на енергия, съхранявана във високоенергийните връзки на молекулата на АТФ.
Активният транспорт на веществата през биологичните мембрани е от голямо значение. Благодарение на активния транспорт в тялото се създават градиенти на концентрация, градиенти на електрически потенциал, градиенти на налягане и др., които поддържат жизнените процеси, тоест от гледна точка на термодинамиката активният транспорт поддържа тялото в неравновесно състояние, осигуряване на нормалното протичане на жизнените процеси.
За извършване на активен пренос, в допълнение към източника на енергия, е необходимо наличието на определени структури. Съгласно съвременните концепции биологичните мембрани съдържат йонни помпи, които работят с енергията на хидролизата на АТФ или така наречените транспортни АТФази, представени от протеинови комплекси.
Понастоящем са известни три типа електрогенни йонни помпи, които активно транспортират йони през мембраната. Това са K + -Na + -ATPase в цитоплазмените мембрани (K + -Na + -помпа), Ca 2+ - ATPase (Ca 2+ -помпа) и H + - ATPase в енергийно-свързващите мембрани на митохондриите (H + - помпа или протонна помпа).
Преносът на йони чрез транспортни АТФази възниква поради свързването на процесите на пренос с химични реакции, дължащи се на енергията на клетъчния метаболизъм.
Когато K + -Na + -ATPase работи, поради енергията, освободена по време на хидролизата на всяка ATP молекула, два калиеви йона се прехвърлят в клетката и три натриеви йона се изпомпват едновременно от клетката. Това създава повишена концентрация на калиеви йони в клетката в сравнение с междуклетъчната среда и намалена концентрация на натрий, което е от голямо физиологично значение.
Благодарение на енергията на хидролизата на АТФ, два калциеви йона се прехвърлят към Ca 2+ -ATPase и два протона се прехвърлят към Н + помпата.
Молекулярният механизъм на действие на йонните АТФази не е напълно разбран. Все пак основните етапи на този сложен ензимен процес могат да бъдат проследени. В случая на K + -Na + -ATPase (нека го обозначим с E за краткост), има седем етапа на йонен трансфер, свързани с хидролизата на АТФ. Обозначенията Е 1 и Е 2 съответстват на местоположението на активния център на ензима върху вътрешната и външната повърхност на мембраната (АДФ-аденозин дифосфат, Р - неорганичен фосфат, звездичката показва активирания комплекс):
1) E + ATP à E*ATP,
2) E*ATP + 3Naà [E*ATP]*Na 3,
3) [E*ATP]*Nа 3 à *Na 3 + ADP,
4) *Na 3 à *Na 3 ,
5) *Na 3 + 2K à *K 2 + 3Na,
6) *K 2 до *K 2,
7) *K 2 à E + P + 2K.
Диаграмата показва, че ключовите етапи на ензима са: 1) образуването на комплекс от ензима с АТФ върху вътрешната повърхност на мембраната (тази реакция се активира от магнезиевите йони); 2) свързване на три натриеви йона от комплекса; 3) фосфорилиране на ензима с образуването на аденозин дифосфат; 4) промяна в конформацията на ензима вътре в мембраната; 5) реакцията на йонен обмен на натрий с калий, протичаща на външната повърхност на мембраната; 6) обратна промяна в конформацията на ензимния комплекс с прехвърлянето на калиеви йони в клетката и 7) връщане на ензима в първоначалното му състояние с освобождаване на калиеви йони и неорганичен фосфат. Така по време на пълен цикъл три натриеви йона се освобождават от клетката, цитоплазмата се обогатява с два калиеви йона и настъпва хидролиза на една молекула АТФ.
В допълнение към йонните помпи, разгледани по-горе, са известни подобни системи, при които натрупването на вещества не е свързано с хидролизата на АТФ, а с работата на редокс ензимите или фотосинтезата. Транспортът на веществата в този случай е вторичен, медииран от мембранен потенциал и (или) градиент на йонна концентрация в присъствието на специфични носители в мембраната. Този транспортен механизъм се нарича вторичен активен транспорт. В плазмените и субклетъчните мембрани на живите клетки е възможно едновременното функциониране на първичен и вторичен активен транспорт. Този механизъм на прехвърляне е особено важен за онези метаболити, за които няма помпи (захари, аминокиселини).
Съвместният еднопосочен транспорт на йони, включващ транспортер от две места, се нарича симпорт. Предполага се, че мембраната може да съдържа носител в комплекс с катион и анион и празен носител. Тъй като мембранният потенциал не се променя в такава схема на трансфер, трансферът може да бъде причинен от разлика в концентрациите на един от йоните. Смята се, че схемата на симпорт се използва за натрупване на аминокиселини в клетките.
Изводи и заключения.
По време на живота клетъчните граници се пресичат от различни вещества, чиито потоци се регулират ефективно. Тази задача се изпълнява от клетъчната мембрана с транспортни системи, вградени в нея, включително йонни помпи, система от молекули-носители и високо селективни йонни канали.
На пръв поглед такова изобилие от системи за пренос изглежда ненужно, тъй като работата само на йонни помпи позволява да се осигурят характерните характеристики на биологичния транспорт: висока селективност, пренос на вещества срещу силите на дифузия и електрическо поле. Парадоксът обаче е, че броят на потоците, които трябва да се регулират, е безкрайно голям, а помпите са само три. В този случай механизмите на йонна конюгация, наречени вторичен активен транспорт, в които дифузионните процеси играят важна роля, стават особено важни. По този начин комбинацията от активен транспорт на вещества с явленията на дифузионен трансфер в клетъчната мембрана е основата, която осигурява жизнената активност на клетката.
Разработено от ръководителя на катедрата по биологична и медицинска физика, кандидат на физико-математическите науки, доцент Новикова Н.Г.
Може също да се интересувате от:

И активен транспорт. Пасивният транспорт се осъществява без консумация на енергия...

ЗАКОНЪТ ФАТИХА И ЖЕНСКИЯТ СУЛТАНТ. Част 2. Исторически факти. Така най-...

Нуждата от реформация и постоянно духовно обновление е Божието изискване за...

Едва ли има период в историята от последните 2500 години, когато определена група от населението...

"Откритията в тази област са свързани с нови технически възможности." Постоянно...

