
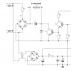





Кръвоснабдяването се осъществява от клоните на две системи - горната и долната артерии (фиг. 19.39). Първият дава клонове: 1) а. ileocolica, която доставя крайния илеум, апендикса, цекума и долната част на илеума.
Ориз. 19,39.Кръвоснабдяване на дебелото черво:
1 - а. mesenterica superior; 2 - а. colica media; 3 - а. colica dextra; 4 - а. илеоколика; 5 - а. mesenter-ica inferior; 6- а. colica sinistra; 7- аа. sigmoideae; 9- а. rektalis superior; 9- а. ректална медия; 70 - а. rektalis inferior
даване; 2) а. colica dextra доставя горната част на възходящото дебело черво, чернодробната кривина и началната част на напречното дебело черво; 3) а. colica media преминава между листата на мезентерията на напречното дебело черво и доставя по-голямата част от това черво (артерията трябва да бъде пощадена по време на операции, включващи дисекция на мезентерията на напречното дебело черво или връзката на стомашно-дебелото черво). В допълнение, връзката на стомашно-дебелото черво, както е показано от проучвания на трупове и наблюдения по време на операции върху пациенти, почти винаги е запоена към мезентерията на напречното дебело черво, главно на нивото на пилоричната част на стомаха. В зоната на сцепление на тези елементи на перитонеума артериалните аркади, образувани от клоните на средната колична артерия, са разположени два пъти по -често, отколкото извън тази зона. Следователно, дисекцията на стомашно-чревния лигамент по време на операции на стомаха трябва да започне 10-12 см вляво от пилора, за да се избегне увреждане на аркадите на средната колична артерия.
Клоните се разклоняват от долната мезентериална артерия: 1) а. colica sinistra, осигуряваща част от напречното дебело черво, изкривяване на далака на дебелото черво и низходящото дебело черво; 2) аа. sigmoideae, отиващи към сигмоидното дебело черво; 3) а. rectalis superior (a. haemorrhoidalis superior - BNA), отивайки в ректума.
Изброените съдове образуват аркади, подобни на тези, открити на тънките черва. Арката, образувана при сливането на клоните на средната и лявата артерии на дебелото черво, преминава между мезентериалните листа на напречното дебело черво и обикновено е добре изразена (преди това се наричаше Риолската арка - arcus Riolani). Той захранва левия край на напречното дебело черво, огъването на далака на дебелото черво и началото на низходящото дебело черво.
При лигиране на горната ректална артерия (във връзка с хирургичното отстраняване на силно локализиран рак на ректума) храненето на първоначалния сегмент на ректума може да бъде рязко нарушено. Това е възможно, защото важен обезпечение, което свързва последната съдова аркада на сигмоидното дебело черво с а, е изключено. haemorrhoidalis (a. rectalis - PNA) superior (вж. фиг. 19.39). Сливането на тази артерия с а. haemorrhoidalis siperior се нарича "критична точка" и се предлага да се лигира ректалната артерия над тази точка - тогава кръвоснабдяването на началния участък на ректума не се нарушава.
Има и други "критични точки" по съдовете на червата. Те включват например багажника a. colica media. Лигирането на тази артерия може да причини некроза на дясната половина на напречното дебело черво, тъй като артериалните аркади а. colica sinistra обикновено не може да осигури кръвоснабдяването на тази част на червата (виж фиг. 19.39).
Крайните форми на разклоняване на долната мезентериална артерия са важни при хирургичното лечение на високо разположени ракови заболявания на ректума, тъй като това изисква мобилизиране на сигмоидното дебело черво с дисекция на мезентерията и лигирането а. haemorrhoidalis superior. Последният представлява крайния клон а. mesenterica inferior. Клиничният опит показва, че такава операция често води до гангрена на частта от ректума, останала след операцията. Същността на въпроса е, че когато лигирате горната ректална артерия, храненето на първоначалния сегмент на ректума може да бъде рязко нарушено. Това е възможно, защото важен обезпечение, което свързва последната съдова аркада на сигмоидното дебело черво с а, е изключено. haemorrhoidalis superior и наречен a. sigmoidea ima. Сливането на тази артерия с a. haemorrhoidalis superior се нарича "критична точка" и се предлага да се лигира ректалната артерия над кръстовището й с посочената колатерала, която най -често се намира на нивото на носа.
А. Ю. Созон-Ярошевич показа, че при хлабава форма на структурата на долната мезентериална артерия могат да се наблюдават повече от един ствол a. haemorrhoidalis superior и два или три ствола, с a. sigmoidea ima в тези случаи е свързан само с един от стволовете на горната ректална артерия. От това следва, че когато артерията се лигира над критичната точка, но под разделянето й на няколко ствола, кръвоснабдяването на частта от ректума ще бъде нарушено.
Изхождайки от това, а също и като вземе предвид други точки (например възможността за вродено отсъствие на долната мезентериална артерия), А. Ю. Созон-Ярошевич предложи да лигира основния й ствол в свободна форма на структурата долна мезентериална артерия. В същото време той вярва, че подобна операция би осигурила по -добре достъп на кръв до крайните клони на долната мезентериална артерия (чрез анастомозите между клоните на горната и долната мезентериална артерия, по -специално чрез a. Colica sinistra). Предложението на А. Ю. Созон-Ярошевич беше успешно реализирано по време на операции на пациенти.
Вените придружават артериите под формата на несдвоени стволове и принадлежат към системата на порталната вена, с изключение на средните и долните вени на ректума, свързани със системата на долната куха вена.
Инервацията на дебелото черво се осъществява от клоните на горния и долния мезентериален сплит. От всички части на червата, най -чувствителната зона към рефлекторни влияния е илеоцекалният ъгъл с апендикса.
Лимфните възли на дебелото черво (nodi lymphatici mesocolici) са разположени по протежение на артериите, които захранват червата. Те могат да бъдат разделени на възли: 1) слепата и апендикса; 2) дебелото черво; 3) ректум.
Възлите на сляпото черво са разположени, както вече споменахме, по клоните на a. ileocolica и нейния ствол. Коловите възли, подобно на мезентериалните, също са подредени в няколко реда. Основните възли на дебелото черво са: 1) на багажника a. colica media, в мезоколона трансверса, до централната група от мезентериални възли; 2) в началото на a. colica sinistra и над него; 3) по ствола на долната мезентериална артерия (виж фиг. 24.17).
19.8. На някои отклонения в структурата и топографията на червата
При изтощени хора, жени, които имат много родители и в напреднала възраст често се наблюдава значителна подвижност на дванадесетопръстника (F.I. Walker).
Сред чревните малформации, срещани на практика, на първо място е дивертикулумът Мека-лъв (дивертикулум Мекели), който съществува при около 2% от хората; това е остатъкът от жлъчно-чревния канал (ductus omphaloentericus), който обикновено прераства до края на 2-рия месец от ембрионалния живот. Дивертикулът е издатина на стената на илеума от страната, противоположна на мезентериума; той е разположен средно на разстояние 50 см от слепото черво (понякога много по -близо до него, понякога по -далеч).
Формата и размерът на дивертикула са много променливи. Най -често има 3 форми на дивертикул: 1) отвор под формата на фистула на пъпа, 2) свързан с пъпа с връв, 3) под формата на сляп джоб на чревната стена.
Възпалението на дивертикулум (дивертикулит) може да бъде сбъркано с апендицит; често дивертикулумът на Мекел е причина за чревна обструкция.
По отношение на дебелото черво трябва да се отбележи, че има редки случаи на ляво възходящо възходящо дебело черво или дясно спускащо се (sinistro и dextropositio coli). Косият ход на напречното дебело черво е по -често срещан, когато flexura coli dextra се намира близо до слепите (което трябва да се има предвид по време на апендектомия), и дългата мезентерия на сигмоидното дебело черво, чиито бримки отиват в дясната половина на коремната кухина (при тази форма на чревна структура може да се наблюдава волвулус) ...
Слепото черво, началният участък на възходящия и крайния участък на илеума понякога имат обща мезентерия - комуна мезентериум ileocaecale, която може да създаде условия за завулвус на цекума.
Вроденото уголемяване на сигмоидното дебело черво (мегасигма), известно като болест на Хиршпрунг, се причинява от рязко намаляване на броя на ганглиозните клетки на сплит Ауербах в дисталното дебело черво. В резултат на това настъпва спастично свиване и стесняване на ректума, което води до вторично рязко разширяване на сигмоидното дебело черво.
Тънките черва се снабдяват с кръв от несдвоените целиакия и черепните мезентериални артерии. Чернодробната артерия, отделена от целиакия, отделя клоните на началната част на дванадесетопръстника. Черепната мезентериална артерия образува дъга по йеюнума, от която много прави артерии, анастомозирани помежду си, се отклоняват към стената на органа.
Тънките черва се инервират от блуждаещия нерв (парасимпатикова нервна система) и постганглионарните клони на лунния ганглий (симпатикова нервна система), които образуват слънчевия сплит.
3. Капиляри: структура и класификация. Органна специфичност на капилярите.
Капиляри
Кръвните капиляри са най -многобройните и тънки съдове. В повечето случаи капилярите образуват мрежи, но те могат да образуват бримки, както и гломерули.
При нормални физиологични условия около половината от капилярите са в полузатворено състояние. Разстоянието между тях е значително намалено, но не го затваря напълно. За формираните елементи на кръвта тези капиляри се оказват непроходими, в същото време кръвната плазма продължава да циркулира през тях. Броят на капилярите в определен орган е свързан с общите му морфофункционални характеристики, а броят на отворените капиляри зависи от интензивността на органа в момента.
Лигавицата на капилярите се формира от ендотела, който лежи върху базалната мембрана. В разцепванията на базалната мембрана на ендотела се разкриват специални разклонени клетки - перицити, които имат множество междинни кръстовища с ендотелни клетки. Отвън капилярите са заобиколени от мрежа от ретикуларни влакна и редки клетки на адвентиция.
Капилярна класификация
Според структурните и функционалните характеристики се разграничават три вида капиляри: соматични, фенестрирани и
Синусоидален или перфориран.
Най -често срещаният тип капиляри е соматичен... В такива капиляри има непрекъсната ендотелна обвивка и непрекъсната базална мембрана. Капилярите от соматичен тип са разположени в мускулите, органите на нервната система, в съединителната тъкан, в екзокринните жлези.
Вторият вид е фенестриранкапиляри. Те се характеризират с тънък ендотел с пори в ендотелните клетки. Порите се затягат от диафрагмата, базалната мембрана е непрекъсната. Фенестрираните капиляри се откриват в ендокринните органи, в чревната лигавица, в кафявата мастна тъкан, в бъбречното тяло и хороидния сплит на мозъка.
Третият тип са капилярите перфориран тип, или синусоида. Това са капиляри с голям диаметър, с големи междуклетъчни и надклетъчни пори (перфорации). Базалната мембрана е прекъсната. Синусоидалните капиляри са характерни за хематопоетичните органи, по -специално за костния мозък, далака, а също и за черния дроб.
Билет 25
1. Цитоплазма. Общи морфологични и функционални характеристики. Класификация на органелите. Структурата и функциите на органелите са от особено значение.
Цитоплазма- вътрешната среда на клетката, затворена между плазмената мембрана и ядрото. Цитоплазмата обединява всички клетъчни структури и улеснява взаимодействието им помежду си.
Това не е хомогенно химично вещество, а сложна, постоянно променяща се физико -химична система, характеризираща се с алкална реакция и високо съдържание на вода.
Всички процеси на клетъчния метаболизъм се извършват в цитоплазмата, с изключение на синтеза на нуклеинови киселини, който се случва в ядрото. прави разлика между два слоя цитоплазма. Външна - ектоплазма Вътрешен слой на цитоплазмата - ендоплазма
Органелисе наричат постоянно присъстващите структури на клетката, които имат определена структура, местоположение и изпълняват определени функции.
Наричат се органели, които постоянно присъстват във всички клетки органели от общо значение.
Други органели присъстват само в някои клетки във връзка с изпълнението на определени специфични функции за тези клетки. Такива органели се наричат органели със специално значение (реснички, микровили, тонофибрили; неврофибрили, миофибрили.)
Органели на цитоплазматаспоред принципа на тяхната структура те се разделят на две групи: мембранаи без мембрана:
· Мембранни органелиса затворени отделения, ограничени от мембрана, която е тяхната стена.
· Немембранни органелине са клетъчни отделения и имат различна структура.
| Реснички и флагели | Те се състоят от 2 части: базално тяло, разположено в цитоплазмата и състоящо се от 9 тройки микротубули и аксонема - израстък над клетъчната повърхност, който е покрит с мембрана отвън, а вътре има 9 двойки микротубули, разположени в кръг , и една двойка в центъра. Между съседни дублети има протеини на нексинов протеин. Радиална спица се простира навътре от всеки дублет. Протеините са прикрепени към микротубулите на централната част, образувайки централна капсула. Динеин протеин, прикрепен към микротубулите (виж по -горе) | Движение на клетката, посока на движение на течността над клетката |
| Микрофиламенти | Тънки нишки, които образуват триизмерна мрежа в клетката. Те се състоят от протеин актин и свързани протеини: фимбрин (свързва паралелни нишки в снопове); алфа-актинин и филамин (свързват нишките, независимо от пространствената им ориентация); винкулин (служи за прикрепване на микрофиламенти към вътрешната повърхност на цитомембраната). Нишките могат да се сглобяват и разглобяват. Миозиновите микрофиламенти, направени от протеина миозин, се намират в малки количества в клетката. Заедно с актина те образуват съкратителни структури | Поддържане на формата на клетката, подкрепа за вътреклетъчни структури, посока на движение на вътреклетъчните процеси, движение и свиване на клетката, образуване на междуклетъчни контакти. Регулиране на клетъчните функции чрез сигнализиране от междуклетъчните контакти за състоянието на извънклетъчния матрикс |
| Микровили са цитоплазмени израстъци с дължина до 1 μm и диаметър 0,1 μm. В сърцевината им има около 40 пролално разположени актинови нишки, те са прикрепени към върха с помощта на протеина винкулин, а в цитоплазмата завършват в крайната мрежа от нишки, където има и миозинови нишки | ||
| Междинни нишки | Дебели здрави нишки с дебелина 8-10 nm, образувани от протеини - виментин, десмин, неврофибриларни протеини, кератин; не може да се монтира самостоятелно-разглобява | Поддържане на клетъчна форма, клетъчна еластичност, участие във формирането на междуклетъчни контакти |
2. Сърдечна мускулна тъкан. Структура и функция. Източници на развитие и регенерация.
PP MT сърдечен (целомичен) тип- развива се от висцерален лист от спланхнатоми, наречен миоепикардна плоча.
В хистогенезата на РР МТ от сърдечен тип се разграничават следните етапи:
1. Етап на кардиомиобластите.
2. Етап на кардиопромиоцитите.
3. Етап на кардиомиоцитите.
Морфофункционалната единица на PP MT от сърдечен тип е кардиомиоцит (CMC). CMC, контактуващи един с друг, образуват функционални мускулни влакна. В същото време самите CMC са разделени един от друг чрез вмъкнати дискове, като специални междуклетъчни контакти. Морфологично CMC е високоспециализирана клетка с едно ядро, локализирано в центъра, миофибрилите заемат основната част от цитоплазмата, между тях има голям брой митохондрии; има EPS и гликогенни включвания. Сарколемата (съответстваща на цитолемата) се състои от плазмолема и базална мембрана, която е по -слабо изразена в сравнение с PP MT от скелетния тип. За разлика от скелетната МТ, сърдечната МТ няма камбиални елементи. В хистогенезата кардиомиобластите са в състояние да се разделят митотично и в същото време да синтезират миофибриларни протеини.
Като се имат предвид характеристиките на развитието на CMC, трябва да се отбележи, че в ранна детска възраст тези клетки, след разглобяване (т.е. изчезване), могат да влязат в цикъла на пролиферация с последващото сглобяване на акто-миозинови структури. Това е характеристика на развитието на клетките на сърдечния мускул. Впоследствие обаче капацитетът за митотично делене в CMC рязко спада и при възрастни е практически нулев. В допълнение, по време на хистогенезата с възрастта, липофусцинови включвания се натрупват в CMC. Размерът на CMC намалява.
Има 3 вида CMC:
1. Свиваема CMC (типична) - вижте описанието по -горе.
2. Атипична (проводима) CMC - образуват проводимата система на сърцето.
3. Секретна CMC.
Атипични (провеждащи CMC - характеризират се с: - слабо развит миофибриларен апарат; - малко митохондрии; - съдържа повече саркоплазма с голям брой гликогенови включвания. Ритъм, способни да произвеждат ритмични нервни импулси, които причиняват свиване на типичния CMC; следователно, дори след прерязване на нервите, приближаващи се към сърцето, миокардът продължава да се свива със собствен ритъм. Друга част от атипичния CMC провежда нервни импулси от пейсмейкъри и импулси от симпатикови и парасимпатикови нервни влакна до контрактилни CMC. Секреторен CMC - разположен в предсърдията ; под електронен микроскоп в цитоплазмата има EPS гранулирани, ламелни комплексни и секреторни гранули, които съдържат натриуретичен фактор или атриопептин - хормон, който регулира кръвното налягане, процеса на уриниране. Освен това, секреторните CMC произвеждат гликопротеини, които, когато се комбинират с липопротеини в кръвта, предотвратяват образуването на кръвни съсиреци в кръвоносните съдове.
Регенерация на PP MT от сърдечен тип.Репаративната регенерация (след увреждане) е много слабо изразена, поради което след увреждане (например: инфаркт) сърдечната МТ се заменя с белег на съединителната тъкан. Физиологичната регенерация (попълване на естественото износване) се осъществява чрез вътреклетъчна регенерация, т.е. CMC не могат да се делят, но постоянно обновяват износените си органели, предимно миофибрили и митохондрии.
3. Далак: структура и функция. Ембрионална и постембрионална хематопоеза.
Далак- хемолимфатичен орган. В ембрионалния период се полага от мезенхимата в началото на 2 -рия месец от развитието. От мезенхимата се образуват капсула, трабекули, ретикуларна тъканна основа, гладкомускулни клетки. Перитонеалната обвивка на органа се формира от висцералния лист на спланнотомите. По време на раждането миелопоезата в далака спира, а лимфоцитопоезата се запазва и засилва.
Структура... Далакът се състои от стромата и паренхима. Стромасе състои от фиброеластична капсула с малък брой миоцити, покрита с мезотелий отвън, и трабекули, простиращи се от капсулата.
V паренхимправи разлика между червена и бяла каша. Червена каша- тя е в основата на орган, изграден от ретикуларна тъкан, пронизан от синусоидални съдове, пълни с кръвни телца, главно еритроцити.Изобилието от еритроцити в синусоидите придава на червената каша червен цвят. Стената на синусоидите е покрита с удължени ендотелни клетки; между тях остават значителни празнини. Ендотелните клетки са разположени върху прекъсната базална мембрана. Наличието на пукнатини в стената на синусоидите прави възможно излизането на еритроцитите от съдовете в околната ретикуларна тъкан. Макрофагите, съдържащи се в големи количества както в ретикуларната тъкан, така и сред ендотелните клетки на синусоидите, увредени от фагоцитоза, стареещи еритроцити, поради което далакът се нарича гробище на еритроцитите. Хемоглобинът на мъртвите еритроцити се доставя чрез макрофаги в черния дроб (протеиновата част - глобинът се използва за синтеза на билирубинов жлъчен пигмент) и червения костен мозък (желязосъдържащ пигмент - хемът се прехвърля в зреещи еритроидни клетки). Друга част от макрофагите участват в клетъчното сътрудничество в хуморалния имунитет (вижте темата "Кръв").
Бяла кашадалакът е представен от лимфни възли. За разлика от възлите на други лимфоидни органи, лимфният възел на далака е проникнат от артерия - а. sentralis. В лимфните възли се разграничават зони:
1. Периартериална зона - е зона, зависима от тимуса.
2. Развъден център-съдържа млади В-лимфобласти (В-зона).
3. Мантийна зона - съдържа предимно В -лимфоцити.
4. Маргинална зона- съотношението на Т- и В-лимфоцити = 1: 1.
По принцип в далака В-лимфоцитите съставляват 60%, Т-лимфоцитите-40%.
Разлики в далака на новородени:
1. Слабо развита капсула и трабекули.
2. Лимфоидната тъкан е дифузна, няма ясни възли
3. В съществуващите лимфни възли центровете на размножаване не са изразени.
Функции на далака:
1. Участие в лимфоцитопоеза (Т- и В-лимфоцитопоеза).
2. Депо за кръв (главно за еритроцити).
3. Елиминиране на повредени, застаряващи червени кръвни клетки
4. Доставчик на желязо за синтеза на хемоглобин, глобин за билирубин.
5. Почистване на кръвта, преминаваща през органа, от антигени.
6. В ембрионалния период - миелопоеза.
Регенерация- много добре, но тактиката на хирурга в случай на наранявания често се определя от особеностите на кръвоснабдяването, поради което е много трудно да се спре паренхимното кървене в органа.
Тираж.Артериалната кръв се насочва към далака през слезната артерия. От артерията има клони, които влизат вътре в големите трабекули и се наричат трабекуларни артерии.От трабекуларната артерия има артерии с малък калибър, които навлизат в червената пулпа и се наричат пулпни артерии. Около пулпните артерии се образуват удължени лимфни обвивки; когато се отдалечават от трабекулата, те се увеличават и придобиват сферична форма (лимфен възел). Вътре в тези лимфни образувания много артерии се отклоняват от артерията, а самата артерия се нарича централна. При излизане от нодула тази артерия се разделя на поредица от клони - четкови артериоли. Овални клъстери от удължени ретикуларни клетки (елипсоиди или ръкави) са разположени около крайните участъци на четките артериоли. В цитоплазмата на ендотела на елипсоидни артериоли се откриват микрофиламенти, които са свързани със способността на елипсоидите да се свиват - функцията на своеобразни сфинктери. По -нататък артериолите се разклоняват в капиляри, някои от тях се вливат във венозните синуси на червената пулпа (теория на затворена циркулация). В съответствие с теорията за отворената циркулация, артериалната кръв от капилярите навлиза в ретикуларната тъкан на пулпата и от нея прониква през стената в синусовата кухина. Венозните синуси заемат значителна част от червената пулпа и могат да имат различни диаметри и форми, в зависимост от кръвоснабдяването им. Тънките стени на венозните синуси са облицовани с прекъсващ ендотел, разположен върху базалната ламина. Ретикуларните влакна преминават по повърхността на синусовата стена под формата на пръстени. В края на синуса, на мястото на прехода му към вената, има друг сфинктер.
В зависимост от намаленото или отпуснато състояние на артериалните и венозните сфинктери, синусите могат да бъдат в различни функционални състояния. Когато венозните сфинктери се свиват, кръвта изпълва синусите, разтяга стената им, докато кръвната плазма преминава през нея в ретикуларната тъкан на пулповите връзки, а кръвните клетки се натрупват в кухината на синусите. Във венозните синуси на далака могат да се задържат до 1/3 от общия брой червени кръвни клетки. Когато и двата сфинктера са отворени, съдържанието на синусите навлиза в кръвта. Често това се случва с рязко увеличаване на нуждите от кислород, когато симпатиковата нервна система се възбуди и сфинктерите се отпуснат. Това се улеснява и от свиването на гладките мускули на капсулата и трабекулите на далака.
Изтичането на венозна кръв от пулпата става през венозната система. Стената на трабекуларните вени се състои само от ендотела, който е в непосредствена близост до съединителната тъкан на трабекулите, тоест тези вени нямат собствена мускулна мембрана. Тази структура на трабекуларните вени улеснява изхвърлянето на кръв от кухината им в вената на далака, която излиза през портата на далака и се влива в порталната вена.
Билет 26
1. Междуклетъчните контакти и тяхната класификация. Синапси. Структура и функция, механизъм на предаване на нервните импулси
ЗАКЛЮЧВАНИЯ
Прост контакт- свързването на клетките поради пръстовидни издатини и издатини на цитомембраните на съседни клетки. Няма специфични структури, които да образуват контакт.
Стегнат контакт- билипидните слоеве на мембраните на съседните клетки са в контакт. В зоната на тесните контакти между клетките практически не преминават вещества.
ЛЕПИЛО
Междуклетъчни адхезионни стави:
Точка- контакт се образува върху малка площ от цитомембрани на съседни клетки.
Адхезивни колани- контактът обгражда цялата клетка по периметъра под формата на колан, намира се в горните участъци на страничните повърхности на епителните клетки.
В контактната зона в цитомембраната са вградени специални трансмембранни протеини, кадхерини, които се свързват с кадхерините на друга клетка.
Калциевите йони са необходими за комбиниране на кадхерини.
От страна на цитоплазмата протеини, бета-катенин, алфа-катенин, гама-катенин, РР-120, ЕВ-1 са прикрепени към кадхерини, а към тях са прикрепени актинови микрофиламенти.
Адхезивни връзки между клетката и извънклетъчния матрикс:
В точката на контакт трансмембранните протеини алфа и бета интегрини са вградени в цитомембраната, които се свързват с елементите на извънклетъчния матрикс.
От страната на цитоплазмата няколко междинни протеини (тензин, талин, алфа-актинин, винкулин, паксилин, фокална адхезионна киназа) са прикрепени към интегрини, към които са прикрепени актинови микрофиламенти.
Десмозоми:
Контактът се формира в малка площ.
В точката на контакт трансмембранните протеини десмоглеин и десмоколин са вградени в цитомембраната, които се свързват със същите протеини на друга клетка.
За да се комбинират десмоколини и десмоглеини, са необходими калциеви йони.
От страната на цитоплазмата междинните протеини - десмоплакин и плактоглобин - са прикрепени към десмоколин и десмоглеин, към които са прикрепени междинни нишки.
ПРОВЕЖДАНЕ
Nexuses (разрязани контакти):
Контактът се формира в малка площ.
На мястото на контакт в цитомембраната са вградени трансмембранни протеини - коннексини, които се свързват помежду си и образуват воден канал в дебелината на мембраната - коннексона.
Връзките на контактуващите клетки са свързани (или съпоставени), в резултат на което между съседните клетки се образува канал, с помощта на който вода, малки молекули и йони, както и електрически ток, свободно преминават от една клетка към друг (в двете посоки).
Синапсът е място, където нервните импулси се предават от една нервна клетка в друга нервна или нервна клетка. В зависимост от локализацията на окончанията на крайните клони на аксона на първия неврон има:
Аксодендритни синапси (импулсът преминава от аксон към дендрит),
Аксосоматични синапси (импулсът преминава от аксона към тялото на нервната клетка),
· Аксоаксонални синапси (импулсът преминава от аксон към аксон).
Според крайния ефект синапсите се разделят: - инхибиторни; - вълнуващо.
Електрически синапс- е натрупване на връзки, предаването се извършва без невротрансмитер, импулсът може да се предава както напред, така и в обратна посока без никакво забавяне.
Химически синапс- предаването се извършва с помощта на невротрансмитер и само в една посока; отнема време за провеждане на импулс чрез химически синапс.
Терминалът на аксона е пресинаптичната част, а областта на втория неврон или друга инервирана клетка, с която контактува, е постсинаптичната част.
Пресинаптичната част съдържа синаптични везикули, множество митохондрии и отделни неврофиламенти. Синаптичните везикули съдържат медиатори: ацетилхолин, норепинефрин, допамин, серотонин, глицин, гама-аминомаслена киселина, серотонин, хистамин, глутамат. Зоната на синаптичен контакт между два неврона се състои от пресинаптичната мембрана, синаптичната цепнатина и постсинаптичната мембрана.
Пресинаптична мембрана- Това е мембраната на клетката, която предава импулса (аксолема). В тази област калциевите канали са локализирани, което улеснява сливането на синаптичните везикули с пресинаптичната мембрана и освобождаването на медиатор в синаптичната цепнатина.
Синаптична цепкамежду пре- и постсинаптичната мембрана е широка 20-30 nm. Мембраните са здраво свързани помежду си в синаптичната област чрез нишки, пресичащи синаптичната цепнатина.
Постсинаптична мембрана- това е част от плазмолемата на клетката, която възприема медиатори и генерира импулс. Той е снабден с рецепторни зони за възприемане на съответния невротрансмитер.
2. Хрущялна тъкан. Класификация, структура и функция. Растеж на хрущяла, неговото регенериране.
Те изпълняват механични, поддържащи, защитни функции. КТ се състои от клетки - хондроцити и хондробласти и голямо количество междуклетъчно хидрофилно вещество, което се характеризира с еластичност и плътност.
Хрущялните клетки са представени от хондробластична диференциация:
1. Стволови клетки
2. Полу-стволови клетки (прехондробласти)
3. Хондробласт
4. Хондроцит
5. Хондрокласт
Стволови и полу-стволови клетки - слабо диференцирани камбиални клетки, локализирани главно около съдовете в перихондриума. Диференцирайки се, те се превръщат в хондробласти и хондроцити, т.е. необходими за регенерация.
Хондробласти - младите клетки са разположени в дълбоките слоеве на перихондриума поотделно, без да образуват изогенни групи. Под светлинен микроскоп хондробластите са сплескани, леко удължени клетки с базофилна цитоплазма.
основната функция на хондробластите- производство на органичната част на междуклетъчното вещество: колагенови и еластинови протеини, глюкозаминогликани (GAG) и протеогликани (PG). В допълнение, хондробластите са способни да се размножават и впоследствие да се превърнат в хондроцити. По принцип хондробластите осигуряват апазиционен (повърхностни, външни неоплазми) растеж на хрущяла от страната на перихондриума.
Хондроцити - основните клетки на хрущялната тъкан, се намират в по -дълбоки слоеве на хрущяла в кухини - лакуни. Хондроцитите могат да се разделят чрез митоза, докато дъщерните клетки не се разминават, те остават заедно - образуват се т. Нар. Изогенни групи. Първоначално те лежат в една обща лакуна, след това между тях и всяка клетка от това се образува междуклетъчно вещество
от изогенната група се появява собствена капсула. Хондроцитите са овално-закръглени клетки с базофилна цитоплазма.
основната функция на хондроцитите- производство на органичната част от междуклетъчното вещество на хрущялната тъкан. Растежът на хрущяла поради разделянето на хондроцитите и производството на междуклетъчно вещество от тях осигурява интерстициален (вътрешен) хрущялен растеж.
В хрущялната тъкан, освен клетките, които образуват междуклетъчното вещество, има и техните антагонисти - разрушители на междуклетъчното вещество - това са хондрокласти(може да се припише на макрофагичната система): доста големи клетки, в цитоплазмата има много лизозоми и митохондрии. Функция на хондрокласт- унищожаване на повредени или износени области на хрущяла.
Междуклетъчно вещество от хрущялна тъкансъдържа колаген, еластични влакна и основно вещество. Основното вещество се състои от тъканна течност и органични вещества: - GAG (хондроетин сулфати, кератосулфати, хиалуронова киселина, липиди. Междуклетъчното вещество е силно хидрофилно, съдържанието на вода достига 75% от масата на хрущяла, което причинява висока плътност и тургор на хрущял. Хрущялната тъкан в дълбоките слоеве няма кръвоносни съдове, храненето се извършва дифузно поради съдовете на перихондриума.
Перихондриуме слоят съединителна тъкан, който покрива повърхността на хрущяла. В перихондриума, външни влакнести(от плътна неоформена КТ с голям брой кръвоносни съдове) слойи вътрешен клетъчен слойсъдържащ голям брой стволови, полу-стволови клетки и хондробласти.
Ембрионална хондрогистогенезаИзточникът на развитие на хрущялната тъкан е мезенхимът.
И. Образуването на хондрогенен примордий или хондрогенен остров.
В някои части на тялото на ембриона, където се образува хрущял, мезенхимните клетки губят своите процеси, интензивно се размножават и плътно прилепнали една към друга, създават определено напрежение - тургор. Стволовите клетки, които са част от островчето, се диференцират в хондробласти. Тези клетки са основните градивни елементи на хрущялната тъкан. В тяхната цитоплазма първо се увеличава броят на свободните рибозоми, след това се появяват участъци от гранулиран ендоплазмен ретикулум.
II. Образуване на първична хрущялна тъкан.
Клетките от централната зона (първични хондроцити) са закръглени, увеличават се по размер, в тяхната цитоплазма се развива гранулиран ендоплазмен ретикулум, с участието на който се осъществява синтезът и секрецията на фибриларни протеини (колаген). Така образуваното междуклетъчно вещество се отличава с оксифилия.
III. Етапи на диференциация на хрущяла.
Хондроцитите придобиват способността да синтезират гликозаминогликани, в допълнение към споменатите по-горе фибриларни протеини, главно сулфатирани (хондроитин сулфати), свързани с неколагенови протеини (протеогликани).
| Тип хрущял | ИНТЕРКЛЕКУЛАТОРНО ВЕЩЕСТВО | Локализация | |
| Фибри | Основно вещество | ||
| хиалинов хрущял | колагенови влакна (колаген II, VI, IX, X, XI) | гликозаминогликани и протеогликани | трахея и бронхи, ставни повърхности, планини, съединения на ребрата с гръдната кост |
| еластичен хрущял | еластични и колагенови влакна | ушна мида, роговиден и клинообразен хрущял на ларинкса, хрущял на носа | |
| влакнест хрущял | успоредни снопове колагенови влакна; съдържанието на влакна е по -голямо, отколкото в други видове хрущяли | места на преход на сухожилия и връзки в хиалинов хрущял, в междупрешленните дискове, полуподвижни стави, симфиза | |
| в междупрешленния диск: фиброзният пръстен е разположен отвън; той съдържа предимно влакна, които имат кръгов ход; а вътре има желатиново ядро - то се състои от гликозаминогликани и протеогликани и хрущялни клетки, плаващи в тях |
Хиалинов хрущял
1. Всъщност в междуклетъчното вещество има голям брой колагенови влакна, чийто коефициент на пречупване е същият като показателя на пречупване на основното вещество; следователно, колагеновите влакна не се виждат под микроскоп; те са прикрити.
2. около изогенни групи има ясно определена базофилна зона - т.нар териториална матрица... Това се дължи на факта, че хондроцитите отделят голямо количество GAGs с кисела реакция, поради което тази област е оцветена с основни бои, т.е. базофилен. Слабо оксифилни области между териториални матрици се наричат междутериториална матрица.
Структурната особеност на хиалиновия хрущял на ставната повърхност е липсата на перихондрий на повърхността, обърната към ставната кухина.
Еластичен хрущял
Особености:
В междуклетъчното вещество, освен колагеновите влакна, има голям брой произволно разположени еластични влакна, което придава еластичност на хрущяла;
· Съдържа много вода;
· Не се калцира (минерални вещества не се отлагат).
Влакнест хрущял
Разположен е в местата на закрепване на сухожилия към костите и хрущялите, в симфизата и междупрешленните дискове. По структура тя заема междинно положение между гъсто оформената съединителна и хрущялна тъкан.
Разлика от другите хрущяли: в междуклетъчното вещество има много повече колагенови влакна, а влакната са ориентирани ориентирано - образуват дебели снопове, ясно видими под микроскоп, постепенно се разхлабват и се превръщат в хиалинов хрущял. Хондроцитите често лежат поотделно по влакната, без да образуват изогенни групи.
Вегетативна инервация на органите
Инервация на окото.В отговор на определени визуални стимули, идващи от ретината, настъпва конвергенция и акомодация на зрителния апарат.
Сближаване на очите- сближаване на зрителните оси на двете очи към разглеждания обект - възниква рефлекторно, с комбинирано свиване на набраздените мускули на очната ябълка. Този рефлекс, необходим за бинокулярното зрение, е свързан с акомодацията на окото. Акомодацията - способността на окото да вижда ясно предмети на различно разстояние от него - зависи от свиването на гладките мускули - m. ciliaris и m. сфинктер зеници. Тъй като дейността на гладките мускули на окото се осъществява заедно със свиването на неговите набраздени мускули, автономната инервация на окото ще се разглежда заедно с животинската инервация на неговия двигателен апарат.
Аферентният път от мускулите на очната ябълка (проприоцептивна чувствителност) е според някои автори самите животински нерви, които инервират тези мускули (III, IV, VI главни нерви), според други - n. офталмикус (n. trigemini).
Центровете на инервация на мускулите на очната ябълка са ядрата на двойки III, IV и VI. Ефективен път - III, IV и VI главни нерви. Сближаването на окото се извършва, както е посочено, чрез комбинираното свиване на мускулите на двете очи.
Трябва да се има предвид, че изолирани движения на една очна ябълка изобщо не съществуват. И двете очи винаги участват във всякакви доброволни и рефлекторни движения. Тази възможност за комбинирано движение на очните ябълки (поглед) се осигурява от специална система от влакна, която свързва ядрата на III, IV и VI нерви и се нарича медиален надлъжен сноп.
Медиалният надлъжен сноп започва в краката на мозъка от ядрото на Даркшевич (виж стр. 503 504), свързва се с ядрата на III, IV, VI нерви с помощта на колатерали и се спуска надолу по мозъчния ствол към гръбначния мозък, където завършва, очевидно, в клетките на предните рога на горните цервикални сегменти. Това позволява движенията на очите да се комбинират с движенията на главата и шията.
Инервация на гладките мускули на окото- м. сфинктер зеници и m. ciliaris, които настаняват окото, възниква поради парасимпатиковата система; инервация на m. dilatator pupillae - поради симпатиковата. Аферентните пътища на автономната система са n. Oculomotorius и n. офталмикус.
Ефективна парасимпатикова инервация Преганглионните влакна идват от ядрото на Якубович (мезенцефалична част от парасимпатиковата нервна система) в състава на n. oculomotorius и по неговия радикс oculomotoria достигат до ганглиозни реснички (фиг. 343), където завършват.
В цилиарния възел започват постганглионните влакна, които чрез nn. ciliares breves достигат до цилиарния мускул и кръговия мускул на ириса. Функция: свиване на зеницата и акомодация на окото за далечно и близко зрение.
Преганглионарните влакна идват от ядрените интермедиолатерални клетки на страничните рога на последния цервикален и два горни гръдни сегмента (CvII - Th11, centrum ciliospinale), излизат през двете горни гръдни rami communicantes albi, преминават през цервикалната част на симпатиковия ствол и завършва в горния цервикален възел. Постганглионните влакна са n. caroticus internus в черепната кухина и влизат в plexus caroticus internus и plexus ophtalmicus; след това част от влакната прониква в ramus communicans, свързвайки се с n. nasociliaris и nervi ciliares longi, а част отива към цилиарния възел, през който преминава без прекъсване в nervi ciliares breves. И тези, и други симпатикови влакна, преминавайки през дългите и късите цилиарни нерви, достигат до радиалния мускул на ириса. Функция: разширяване на зеницата, както и стесняване на съдовете на окото.
Инервация на слъзните и слюнчените жлези.Аферентният път за слъзната жлеза е n. lacrimalis (клон на n. ophthalmicus от n. trigemini), за подчелюстната и сублингвалната - n. Iingualis (клон n. Mandibularis от n. Trigemini) и chorda tympani (клон n. Intermedins), за паротид - n. auriculotemporalis и n. glossopharyngeus.
Ефективна парасимпатикова инервация на слъзната жлеза... Центърът се намира в горната част на продълговатия мозък и е свързан с ядрото на междинния нерв (nucleus salivatorius superior). Преганглионните влакна са n. intermedius, оттук нататък n. petrosus major до ганглий птеригопалатин (фиг. 344).

От тук започват постганглионните влакна, които са част от n. maxillaris и по -нататък неговите клони n. зигоматика чрез връзки с n. lacrimalis достигат до слъзната жлеза.
Ефективна парасимпатикова инервация на субмандибуларните и сублингвалните жлези... Преганглионните влакна идват от ядрото slivatorius superior като част от n. intermedius, по -нататък chorda tympani и n. lingualis до ганглий субмандибуларен, откъдето започват постганглионарните влакна, достигащи до жлезите в езиковия нерв.
Ефективна парасимпатикова инервация на паротидната жлеза.Преганглионните влакна идват от ядрото slivatorius inferior като част от n. glossopharyngeus, по -нататък n. tympanicus, n. petrosus minor към ganglion oticum (фиг. 345).

От тук започват постганглионарни влакна, отиващи към жлезата като част от n. auriculotemporalis. Функция: засилване на секрецията на слъзните и слюнчените жлези; вазодилатация на жлезите.
Ефективна симпатична инервация на всички тези жлези.Преганглионарните влакна започват в страничните рога на горните гръдни сегменти на гръбначния мозък и завършват в горния шиен възел. Постганглионарните влакна започват в посочения възел и достигат до слъзната жлеза като част от plexus caroticus internus, до паротида - като част от plexus caroticus externus и до подмандибуларните и сублингвалните жлези - през plexus caroticus externus и след това през plexus facialis . Функция: забавяне на отделянето на слюнка (сухота в устата). Лакримация (ефектът не е остър).
Инервация на сърцето(фиг. 346).

Различни пътища от сърцето са n. vagus, както и в средните и долните цервикални и гръдни сърдечни симпатикови нерви. В този случай усещането за болка се осъществява по симпатиковите нерви, а всички други аферентни импулси се извършват по протежение на парасимпатиковите нерви.
Преганглионните влакна започват в дорзалното вегетативно ядро на блуждаещия нерв и са част от последния, неговите сърдечни разклонения (rami cardiaci n. Vagi) и сърдечни сплитания към вътрешните възли на сърцето, както и възлите на перикардните полета. Постганглионните влакна излъчват от тези възли към сърдечния мускул. Функция: инхибиране и потискане на сърдечната дейност. Стесняване на коронарните артерии.
Ако Цион през 1866 г. откри "нервно-чувствителния" нерв, който върви центростремително като част от блуждаещия нерв. Намаляването на кръвното налягане е свързано с този нерв, поради което е кръстен n. депресор.
Ефективна симпатична инервация.Преганглионарните влакна започват от страничните рога на гръбначния мозък 4-5 горни гръдни сегмента, излизат като част от съответните rami communicantes albi и преминават през симпатиковия ствол до пет горни гръдни и три шийни възли. В тези възли започват постганглионните влакна, които са част от сърдечните нерви, nn. cardiaci, cervicales superior, medius et inferior и nn. cardiaci thoracici, достигат до сърдечния мускул. Според К. М. Биков и др., Прекъсването се извършва само в ганглийния стелатум. Според описанието на Г. Ф. Иванов, сърдечните нерви съдържат преганглионни влакна, които преминават към постганглионарни в клетките на сърдечния сплит. Функция: укрепване на работата на сърцето и ускоряване на ритъма, разширяване на коронарните съдове.
Инервация на белите дробове и бронхите.Аферентните пътища от висцералната плевра са белодробните клони на гръдния симпатиков ствол, от париеталната плевра - nn. intercostales и n. phrenicus, от бронхите - n. блуждаещ.
Ефективна парасимпатикова инервация.Преганглионните влакна започват в дорзалното автономно ядро на блуждаещия нерв и отиват като част от последния и неговите белодробни клони към възлите на plexus pulmonalis, както и към възлите, разположени по протежение на трахеята, бронхите и вътре в белите дробове. Постганглионните влакна са насочени от тези възли към мускулите и жлезите на бронхиалното дърво. Функция: стесняване на лумена на бронхите и бронхиолите и отделяне на слуз; вазодилатация.
Ефективна симпатична инервация.Преганглионарните влакна излизат от страничните рога на гръбначния мозък на горните гръдни сегменти (Th2-Th6) и преминават през съответните rami communicantes albi и симпатиковия ствол към звездните и горните гръдни възли. От последното започват постганглионарни влакна, които преминават като част от белодробния сплит към бронхиалните мускули и кръвоносните съдове. Функция: разширяване на лумена на бронхите. Свиване и понякога разширяване на кръвоносните съдове.
Инервация на стомашно -чревния тракт (до сигмоидното дебело черво), панкреаса, черния дроб.Различни пътища от тези органи са n. vagus, n. splanchnicus major et minor, plexus hepaticus, plexus celiacus, гръдни и лумбални гръбначни нерви, и според F.P. Polyakin и I.I. Shapiro, и като част от n. phrenicus.
Усещането за болка от тези органи се предава по симпатиковите нерви, по n. vagus - други аферентни импулси, а от стомаха - чувство на гадене и глад.
Ефективна парасимпатикова инервация.Преганглионните влакна от дорзалното автономно ядро на блуждаещия нерв преминават като част от последния до крайните възли, разположени в дебелината на тези органи. В червата това са клетки на чревния сплит (plexus myentericus, submucosus). Постганглионните влакна се простират от тези възли до гладки мускули и жлези. Функция: укрепване на перисталтиката на стомаха, отпускане на сфинктера на пилора, увеличаване на перисталтиката на червата и жлъчния мехур. По отношение на секрецията, блуждаещият нерв съдържа влакна, които го възбуждат и инхибират. Разширяване на кръвоносните съдове.
Ефективна симпатична инервация.Преганглионарните влакна излизат от страничните рога на гръбначния мозък на V-XII гръдните сегменти, преминават по съответния rami communicantes albi в симпатиковия ствол и след това без прекъсване в nn. splanchnici majores (VI-IX) до междинни възли, участващи в образуването на слънчевия и долния мезентериален сплит (ganglia celiaca и ganglion mesentericum superius et inferius). Оттук възникват постганглионни влакна, които са част от сплит celiacus и pi. tesentericus превъзхождащ черния дроб, панкреаса, тънките черва и дебелото черво до средата на напречната част; лявата половина на дебелото черво transversum и colon descendens са инервирани от plexus mesentericus inferior. Тези сплетения захранват мускулите и жлезите на тези органи. Функция: забавяне на перисталтиката на стомаха, червата и жлъчния мехур, стесняване на лумена на кръвоносните съдове и инхибиране на секрецията на жлезите.
Към това трябва да се добави, че забавянето на движенията в стомаха и червата се постига и от факта, че симпатиковите нерви предизвикват активно свиване на сфинктерите: сфинктер пилори, чревни сфинктери и др.
Инервация на сигмоидното дебело черво, ректума и пикочния мехур... Различни пътища преминават като част от plexus mesentericus inferior, plexus hypogastrics superior и inferior и като част от nn. splanchnici pelvini.
Ефективна парасимпатикова инервация. Преганглионните влакна започват в страничните рога на гръбначния мозък на II-IV сакрални сегменти и излизат като част от съответните предни корени на гръбначните нерви. Тогава те идват под формата nn. splanch-nici pelvini към вътрешноорганични възли на посочените участъци на дебелото черво и околоорганни възли на пикочния мехур. В тези възли започват постганглионарни влакна, които достигат до гладката мускулатура на тези органи. Функция: стимулиране на перисталтиката на сигмоида и ректума, отпускане на m. сфинктер ani internus, свиване на m. detrusor urinae и релаксация T. sphincter vesicae.
Ефективна симпатична инервация.Преганглионните влакна преминават от страничните рога на лумбалния гръбначен мозък през съответните предни корени в rami communicantes albi, преминават без прекъсване през симпатиковия ствол и достигат до ganglion mesentericum inferius. Тук започват постганглионните влакна, които са част от nn. hypogastrici до гладките мускули на тези органи. Функция: забавяне на перисталтиката на сигмоида и ректума и свиване на вътрешния сфинктер на ректума. В пикочния мехур симпатиковите нерви причиняват отпускане на m. детрузорни урини и свиване на сфинктера на пикочния мехур.
Инервация на гениталиите: симпатичен, парасимпатичен. Инервацията на други вътрешни органи е дадена след тяхното описание.
Инервация на кръвоносните съдове.Степента на инервация на артериите, капилярите и вените не е еднаква. Артериите с по -развити мускулни елементи в туниката получават по -обилна инервация, вените по -малко; v. cava inferior и v. portae са между тях.
По -големите съдове, разположени вътре в телесните кухини, получават инервация от клоните на симпатиковия ствол, най -близките сплитания на автономната система и съседните гръбначни нерви; периферните съдове на стените на кухините и съдовете на крайниците получават инервация от нервите, преминаващи наблизо. Нервите, приближаващи се до съдовете, преминават сегментарно и образуват периваскуларни сплетения, от които влакната проникват през стената и се разпределят в адвентицията (tunica externa) и между последната и туниката. Влакната доставят мускулните образувания на стената, имащи различни окончания. В момента наличието на рецептори е доказано във всички кръвоносни и лимфни съдове.
Първият неврон на аферентния път на съдовата система се намира в междупрешленните възли или възлите на автономните нерви (nn. Splanchnici, n. Vagus); след това отива като част от проводника на интероцептивния анализатор. Вазомоторният център се намира в продълговатия мозък. Палиусът на глобуса, зрителният туберкул, а също и сивият туберкул са свързани с регулирането на кръвообращението. Висшите центрове на кръвообращение, както всички автономни функции, се намират в кората на двигателната зона на мозъка (челен лоб), както и отпред и зад него. Според най -новите данни кортикалният край на анализатора на съдови функции се намира, очевидно, във всички части на кората. Низходящите връзки на мозъка със стволови и гръбначни центрове се осъществяват, очевидно, от пирамидалните и екстрапирамидалните пътища.
Затварянето на рефлекторната дъга може да се случи на всички нива на централната нервна система, както и във възлите на автономния сплит (собствена автономна рефлекторна дъга).
Еферентният път причинява вазомоторен ефект - вазодилатация или вазоконстрикция. Вазоконстрикторните влакна са част от симпатиковите нерви, съдоразширяващите влакна са част от всички парасимпатикови нерви на черепната част на автономната система (III, VII, IX, X), като част от задните корени на гръбначните нерви (не разпознат от всички) и парасимпатикови нерви на сакралната част (nn. Splanchnici pelvini).
Лекция 30
ЧРЕВНА ИНЕРВАЦИЯ. - ДЕФЕКТ. - ИЗСИМВАНЕ, ТЕХНИКА НА ИЗСЛЕДВАНЕТО. - ИЗСУКВАНЕ НА СОЛНИ РАЗТВОРИ И КРЪВЕН СЕРУМ. - НАЧИНИ НА ВСАСВАНЕ
Преди това зависимостта на движението на червата от нервите се разглежда по такъв начин, че блуждаещият нерв се счита за двигателен нерв, а n. забавяне на splanchnicus. Сега въпросът за инервацията на червата стана изключително сложен, но като цяло все още се смята, че блуждаещият нерв е двигателният нерв, а n. splanchnicus - Задържащ нерв. Що се отнася до подробната настройка на експериментите, подробности, следва да се отбележи следното. Ако директно дразните блуждаещия нерв, тогава често няма да забележите появата на движение на червата при животното или ще получите нещо неясно, неясно. Опитът става по -добър, ако първо отрежете n. splanchnicus, т.е. симпатиковия нерв. Тогава действието на вагуса става по -изразено. Какво означава? И това трябва да се разбира по този начин. При гладно животно, което не смила нищо, храносмилателният канал е в покой. Тази почивка се дължи на действието на забавящия нерв.
Следователно, ако раздразните блуждаещото животно в гладно животно, в което действат забавящите нерви, тогава ще срещнете антагонистично действие от страна на n. splanchnicus. Оказва се „борба“ с нерви, а цялостната картина, крайните резултати стават несигурни. Следователно, за да се получи ясно изразена възбуда на червата при раздразнение на вагуса, първо е необходимо да се отървете от влиянието на забавящите нервите. Този факт трябва да ви напомни за друг факт, който вече съобщих, а именно по отношение на отделянето на чревен сок. Там казах, че единственият известен факт е, че след прекъсването на мезентериалните нерви има непрекъснато отделяне на чревния сок. Това последно явление трябва да бъде разбрано по такъв начин, че от нервите да произтича забавящо влияние; когато ги нарежете, тогава отделянето на сока продължава без забавяне и става много обилно.
Това означава, че в този случай имаме факт, подобен на предишните. И тук се оказва, че постоянното действие на нервите се забавя.
Следователно, както по отношение на секрецията на червата, така и във връзка с тяхното движение, виждаме малко по -различен план за нормалната активност на нервите. Тук действието на нервите се забавя, не е възбуждащо, не е същото като например върху скелетните мускули. При съществуването на забавяща функция n. следователно splanchnicus може да се разглежда и в положителна форма. Ако има движения в червата, причинени или от дразнене на нервите, или по друг начин, тогава дразнене n. splanchnicus ще спре тези движения. Следователно действие n. splanchnicus се доказва по два начина: както чрез спиране на движението на червата, когато е раздразнено, така и чрез появата на отчетливи движения след разрязване, когато вагусът е раздразнен.
Както можете да видите, тази глава за изхождането е много по -кратка от предишните. Не е по -просто, но тук има по -малко факти. Факт е, че много въпроси далеч не са уредени тук, но оскъдността на фактите зависи от факта, че физиолозите са изучавали тази област малко и не според правилния план.
За да ми остане да кажа повече за онези факти, свързани с изхвърлянето на остатъци от храна, за дефекация, екскременти. Това се случва на дълги интервали, което става възможно, защото има специални ключалки, сфинктери. Сфинктерите се инервират от специални нерви, под въздействието на специална нервна система, освен това под влияние на два вида нерви: забавящи и възбуждащи.
Когато настъпи актът на дефекация, сфинктерите се дразнят, което води до тяхното отпускане и до отваряне на ануса. И когато трябва да се предотврати дефекацията, настъпва силно свиване на сфинктерите. Нервните влакна, които инервират сфинктерите, отиват в n. hypogastricus и n. еригени.
Актът на дефекация е рефлексен. Необходимостта от движение на червата се усеща чрез сензорните нерви, разпръснати в ректума. Що се отнася до центровете, благодарение на които се осъществява рефлексът, има няколко от тях: в долната част на червата, в гръбначния мозък и дори в мозъка. Следователно институциите на нервните центрове са разположени на няколко етажа. Това може да се провери чрез клинични данни, лабораторни наблюдения и личен опит. Трябва първо да се разпознаят най -близките центрове на самото черво, след това центровете в гръбначния мозък и накрая центровете в мозъчните полукълба. Долните центрове са изградени от ганглии в коремната кухина. Че съществуват такива центрове и те трябва да бъдат разпознати, това се доказва от факта, че ако при едно животно целият гръбначен мозък е унищожен, започвайки от първите гръдни или дори цервикални части, то при такова животно без гръбначен мозък, пълна първоначално се наблюдава нарушение на отделителния механизъм, но постепенно всичко приема нормален характер. Очевидно е намерен управленски апарат за сфинктерите, открити са центрове. Те трябва да се имат предвид в долните центрове на коремната кухина.
Нека сега се включим в опита. Пред нас е заек, отровен с хлоралхидрат. Коремната му кухина беше отворена и n. vagus се взема с лигатура. Когато вагусът е раздразнен, движенията на червата са видими и махалото и перисталтично. Експериментът не беше напълно успешен, тъй като n не беше отрязан. splanchnicus и резултатът беше борба между забавящите и двигателните нерви. Продължавам по отношение на инервацията на сфинктерите на ануса. И така, първата инервация, където се извършва прехвърлянето на центростремителни стимули към центробежни, е в някакъв ганглий извън централната нервна система. Следващият случай е лумбалната област. Това е лесно да се провери както при животни, така и при клинични наблюдения. Клиницистите са наясно с факта, че при заболявания на гръбначния мозък човек често прави дефекация против волята си. Ако гръбначната част на мозъка е унищожена при животно, тогава се получава и нарушение в акта на дефекация.
Тогава, както знаем от собствен опит, последната, най -висша инстанция на нервните центрове достига до мозъчните полукълба. При хората и в масата на животните актът на екскременти е напълно произволен. Това служи като доказателство, че рефлекторната дъга може да бъде затворена и през мозъчните полукълба.
Така че, за такъв на пръв поглед прост въпрос като дефекацията, има, както видяхме, такъв сложен рефлексен акт. С това ще завърша представянето си на въпроса за двигателната работа на храносмилателния канал.
Сега се обръщам към третата работа на храносмилателния апарат - абсорбционната работа. Процесът на усвояване е тясно свързан с работата на храносмилането и движението. Благодарение на храносмилането храната се опростява по химичния си състав и благодарение на движенията на червата, тя се размазва, движи се по целия храносмилателен канал. Всичко това има за цел да направи храната усвоима. Докато хранителните вещества, които сме приели, остават в стомаха и червата, те са външни вещества за организма и лесно могат да бъдат отстранени от него. Едва когато отидат по -дълбоко, отвъд стените на червата, те стават собственост на тялото.
По отношение на усвояването много учени отдавна започнаха да изразяват мнението си, но този въпрос все още не е напълно изяснен и представлява един вид кост на раздора между физиолозите.
Знаете от физиката, че през пропускливи и полупропускливи мембрани веществата преминават от един съд в друг. Това са така наречените дифузионни и осмотични явления. И така, когато физиолозите стигнаха до процеса на усвояване, те вярваха, че ситуацията тук е проста: преминаването на преработената храна през стените на червата е същото като през мъртвите мембрани. Както вече трябва да знаете за химията на храносмилането, цялото съдържание на храносмилателния тракт, поне това, което тялото ще усвои за собствените си цели, влиза в разтвор. Крайната цел на химията е да превърне всичко в разтворени, лесно дифузни вещества. Естествено, физиолозите смятат, че по -нататък, когато храносмилането приключи, има просто преминаване на храната през стените на червата във вътрешността на тялото. Въпросът обаче се оказа не толкова прост.
Да, ето една малка бележка. Предоставих ви фактите, свързани със секреторната и двигателната активност на храносмилателните органи и преминах към абсорбционната дейност. Но пропуснах един раздел, тези, които внимателно ме слушаха, можеха да го забележат. Това е отделът, който се занимава с подробна химия на храносмилането. След като говорих за ензимите, за това как те работят, би било необходимо да се проучи, за да се разбере как всъщност се извършва обработката на входящите вещества в храносмилателния канал. Например, колко протеини, мазнини, въглехидрати се усвояват във всеки отдел, какви продукти на разпадане могат да бъдат открити тук и там и т. Н. Тези факти, разбира се, са много интересни и имат пряко отношение към това, което ви чета, но ги пропускам, тъй като те принадлежат към областта на физиологичната химия. Всичко това, разбира се, е една и съща физиология, но темата е нараснала много и от удобство физиологът ви разказва за едно, а химикът-физиолог за другото. Всичко това ще ви бъде съобщено своевременно и аз ще премина към нещата, които засягат моя отдел.
Това означава, че абсорбцията е прехвърляне на подготвени вещества в дълбините на тялото за смесване със соковете на тялото и за влизане в състава на живата материя.
Първоначално те бяха склонни да разглеждат този преход като явление на осмозата. Вярно е, че това беше още през четиридесетте и петдесетте години на миналия век. До тридесетте и четиридесетте години физиологията беше доминирана от концепция, която беше много вредна и антинаучна, а именно, те мислеха за някаква специална „жизнена сила“. Това беше така нареченият витализъм. За всичко, каквото и да се случи в животинския организъм на непонятното, отговорът беше един и същ, че това е „жизнената сила“, която го прави. Тази дума по онова време обясняваше всичко и премахваше всякаква нужда от строго научно обяснение. Ясно е, че този витализъм само затвори пътя за истински научни изследвания, които свеждат сложните явления до по -прости, вече установени или от дадена наука - физиология, или от други науки: механика, физика, химия и т. Н. Когато физиолозите осъзнаха, че „Жизнена сила“ е празна дума, безполезна и не обясняваща нищо, след което те започнаха да свеждат всички явления на живота, всички физиологични факти до физически и химически явления. Задачата на физиологичните изследвания беше да обяснят всичко чрез физични и химични закони. Това се разглеждаше като истинско научно предизвикателство. Физиолозите скочиха на нова идея. По това време бяха предложени физико -химични обяснения за много процеси. За много груби явления тези обяснения се оказаха много подходящи. За по -фина физиология, например, за живота на клетката, тези обяснения не пасват и скоро са изоставени и забравени. Това е разбираемо. Храносмилателната, например, активност, както можете да видите, е истинска химическа активност, която трябва да се изучава с чисто химични методи. Същото, както ще видите по -нататък, може да се каже за кръвообращението, за работата на сърцето. Там се случват чисто физически процеси. Идеята за сърцето, грубата идея за помпа, е добра. Всички физико -химични обяснения, приложени към големи части, към цели органи, се оказаха доста успешни и приемливи, докато тези, приложени към фини части, към клетката, се оказаха неправилни и всички впоследствие изчезнаха. Това се обяснява с факта, че знаем повече за дейността на голям орган, по -лесно е да го изучаваме и по -лесно се доближаваме до макроскопичен орган. Активността на клетката е почти напълно неизвестна за нас. Ясно е, че обясненията за работата на онези органи, които познаваме, се оказаха подходящи, а обясненията на това, което не знаем, бяха неуместни.
И така, абсорбцията през чревната стена изглеждаше първоначално прост акт, тя се разглеждаше като проста осмоза. Но когато се запознахме по -отблизо с темата, се разкри голямо разминаване между това, което физикът даде за разбиране и това, което е в действителност. Сега има отклонения от чисто физически обяснения по цялата линия. Въпросът трябва да бъде разбран по такъв начин, че законът все още да не се вижда зад детайлите, които излязоха. Разбира се, нито един от физическите и химичните закони не е нарушен от живо същество. Но освен физико -химичните, съществуват свои собствени закони, много сложни и ние все още не ги разбираме, те са обхванати от маса детайли, детайли, чието значение не ни е напълно ясно.
Тенденцията на физиолозите да сведат цялата активност на организма до физични и химични закони, да дадат на всичко физическо обяснение, в крайна сметка предизвика реакция. Това винаги се случва с едностранна страст към нещо. Тази реакция, този обрат в науката се нарича неовитализъм, нов витализъм. Всъщност възкресението на витализма означава само, че физико-химичното обяснение, което процъфтява в края на петдесетте, дава много лоши тълкувания и се оказва неприложимо за клетъчната физиология. Тогава противоположното мнение вдигна глава. Но това означава само, че все още не знаем всичко, че все още не са разработени средства за извършване на строго научен анализ на живота на клетката, за продължаване на въпроса, както вече го правим с големи органи. И, разбира се, появата на неовитализма не може да бъде разбрана така, сякаш сме изгонили „жизнената сила” от големите органи, но тя остана в малките. Той само показва състоянието на нашите знания. Клетъчната физиология тепърва започва да се развива, получават се само първите фрагментарни факти. Известно е, че имаше време, когато дейността на големите органи изглеждаше мистериозна и не отговаряше на физико -химичното разбиране. И сега ние работим изключително с тези понятия и не въвеждаме други. Сега цялата „мистерия“ е открита и повторена в нашите чаши. Изучавате редица ензими и химическата им работа се извършва в епруветки пред очите ви.
Това ще отнеме 10-20 години и всички ензими ще бъдат изследвани от гледна точка на тяхната химическа природа. Клетъчната физиология също ще напредне. Така трябва да се разбират онези случаи, когато физико -химичните обяснения не са приложими в момента. Това означава, че още не е дошъл моментът, че още не знаем всичко. Разбираемо, трябва да се отдаде заслуга на премахването на жалките обяснения. Често в науката има един вид измама на сетивата - изглежда, че разбирате, но всъщност не разбирате. Това се случи и с физико-химичните познания. Това, разбира се, не е добродетел, а порок, такава самозаблуда замъглява истината. Следователно, когато истински учен отхвърля лошите обяснения на явления, дори тези обяснения да са физико -химични, това не е триумф на неовитализма, а само строго отношение към обяснението. Това изобщо не изключва възможността да се намери правилния, солиден и напълно научен път, който да поеме нещата в бъдеще, както се е случвало повече от веднъж в миналото. И така, физиолозите от четиридесетте години смятат, че абсорбцията е прост осмотичен процес. Но тогава физиологът Хайденхайн опроверга тази позиция. Той представи факти, които противоречат и унищожиха физико -химичните обяснения. Борбата със старите физиологични концепции е много поучителна. Не е безполезно да се спираме на това.
Следователно абсорбцията се разглежда по -рано като прост осмотичен процес, насочен към изравняване на състава на веществото от едната и от другата страна на чревните стени. Абсорбцията води до уравнение на състава. Всичко това са чисто физически представи. Във физиката, както знаете, има подробна теория за осмотичните явления, теорията на Вант Хоф. Van't Hoff третира разтворените твърди вещества като газове. Газовете обикновено са равномерно разпределени. Това означава, че в този случай, в случай на разтворени вещества, ако имате мембрана между тях, веществата ще се разпределят равномерно от двете му страни. Но за това е необходимо да има разлика в състава, едва тогава ще започне изравняването. Ако няма такава разлика в композицията, тогава движението няма да започне, няма да има нужда от това.
Нека се обърнем към абсорбцията. Всичко, което се намира в храносмилателния канал, всичко се превежда в лимфата, в кръвта, с една дума, в соковете на тялото. Следователно, за да може да се говори за осмотични явления, трябва да има разлика в състава. Но какво се оказва? Самият факт, че каквото и да влезете в храносмилателния канал, след това преминава в соковете на тялото, само този факт показва, че преходът на веществата става независимо от състава на приеманата храна. И Хайденхайн доказа в редица експерименти, че в този случай физикохимичното обяснение не отговаря на явлението, не го обхваща напълно.
Тези експерименти са както следва. Нека вземем разтвор на готварска сол. Както ви казах, основната телесна течност е 0,9% разтвор на натриев хлорид. Тази течност измива цялото тяло. Ако премахнем всички формирани елементи, протеини и т.н. от нашите сокове, тогава ще остане само вода, този 0,9% разтвор на сол. Следователно такова решение се нарича физиологично. Така че, изглежда така; ако излеете 0,9% разтвор на натриев хлорид в храносмилателния канал, тогава той не трябва да излиза извън стените, тъй като готварската сол е в него в същото съотношение, както е в тялото. Ще получите изотоничен разтвор с телесни сокове, със същия химичен тон. Оказва се обаче, че това решение влиза в тялото и не остава в червата.
Можете да отидете по -далеч. Можете да вземете кръвен серум, тоест тази течност, която прониква във всичко, в цялото тяло (разбира се, с изключение на морфологичните елементи, които не се броят). И този серум, въведен в храносмилателния канал, също излиза от него в тялото. Това означава, че макар да няма основно условие за проста осмотична абсорбция, преходът, между другото, се осъществява.
Сега ще направим този експеримент на Хайденхайн. Имаме куче, при което коремната кухина се отваря и част от червата се изолира по време на прехода от дванадесетопръстника към йеюнума за 40 см. В този изолиран участък ще въведем изотоничен разтвор на натриев хлорид, тоест физиологичен разтвор. Следователно, според осмотичните закони, не трябва да има движение на този разтвор вътре в тялото. Но ще видите, че този изотоничен разтвор ще отиде в другата страна на червата. Ако имахме физическо устройство, разделено от мембрана, тогава при такива условия решенията биха останали неподвижни. И така, ще изсипем 80 кубически метра в празното черво. см физиологичен разтвор и след 15 минути ще видим какво ще се случи.
Сега въпросът е: какво се случва, ако се инжектират неизотонични разтвори? Ако например се излее хипертоничен или хипотоничен разтвор, т.е. съдържащ повече или по -малко сол от телесната течност, тогава според осмотичната теория би трябвало да се очаква следното. Ако това е 2% разтвор на сол, тогава трябва да се очаква, че водата от тялото ще отиде към солта, в червата, ще се получи увеличаване на излятия разтвор и това ще изравни съставите. И ако имате 0,5% или 0,3% разтвор, тогава трябва да очаквате, че водата първо ще напусне червата, за да направи разтвора в червата по -концентриран. Не се случва нито едното, нито другото. Всички разтвори вървят по същия начин и преминават от другата страна на червата. Няма съответствие с това, което човек би очаквал. Но не е необходимо, разбира се, да се разбере, че това е нарушение на осмотичния закон. Това не е така. Има само усложнение на явлението; когато изучите добре всички подробности, ще намерите този закон и тук.
Хайденхайн добави към това преживяване. Той се опита да отнеме от стените на червата жизнените им свойства, живата им природа. Той постигна това чрез инжектиране на вещества като натриев флуорид в храносмилателния канал, който действа убийствено върху тъканите, отнемайки жизнените им свойства. И тогава в червата беше точно същото като в стъклото на физика. Тогава изотоничният разтвор не премина, но хипертоничният и хипотоничният разтвор преминаха през червата. По този начин, веднага щом сложните свойства на живата чревна мембрана бяха унищожени, действията на физическите закони веднага бяха ясно разкрити. Следователно живата стена варира в своята дейност, затъмнявайки действието на физическите закони.
Когато Хайденхайн публикува неговите произведения, неовиталистите го включиха до известна степен в своя „полк“. Те си представяха, че той застъпва неовиталистична гледна точка. За Хайденхайн това, разбира се, беше обида. Болеше го. Има и една много интересна статия от Хайденхайн, където той очертава отношението си към тази гледна точка: едно е, посочи той, да се вземат предвид физическите обяснения на всички факти, които винаги са на разположение, и друго е да се считат всички явления за никога научно обяснимо. И накрая, все още можем да приемем, че физическите обяснения, които не са налични днес, ще станат достъпни с течение на времето. Да се разбере и обясни всичко е идеалът за науката.
Нека се върнем към първия ни опит. Сега същият заек има nn. splanchnici се нарязват и лигират. Дишането е изкуствено. Абсолютна почивка на червата. Дразни блуждаещия. Червата започнаха да се движат. За съжаление трябва само да слушате, а не да виждате. Докато nn. splanchnici бяха непокътнати, можехме да предизвикаме движение само за кратко време, но сега действието на вагуса е доста отчетливо.
Забавящият нерв вече е изчезнал и след като раздразнихме вагуса, получихме движение, което не спираше дълго време и с последващи раздразнения само засилихме предишното движение. Тези движения наподобяват капризите на куп червеи. Тук се забелязват предимно движения, подобни на махало. Тъй като тези движения не спират, ще покажем действието на забавящия нерв. Ще го хванем и ще го дразним. Досадно. Все още има движение. Не се забелязват ясни действия. За да има пълно забавяне, и двата nn трябва да се дразнят. splanchnici. Във всеки случай има следния факт. Докато nn не бяха отрязани. splanchnici, имахме пълна, стабилна почивка. Сега, напротив, не можем да спрем да се движим. Под влияние на дразнене на вагуса се наблюдава увеличаване на движението. Следователно, по отношение на забавящите нерви, ние изхождаме от факта, че сме получили.
Фактът за съществуването на забавящ нерв поражда съмнения сред много изследователи. Спорът е решен чрез намаляване на nn. splanchnici; след това имаше рязко действие на вагуса - двигателния нерв. Ето една аналогия с действието на вагуса по отношение на панкреатичната жлеза.
Нека се обърнем към втория ни експеримент. Въведени са 80 кубически метра. вижте физиологичен разтвор на натриев хлорид. Да видим какво се е случило. Изминаха 15 минути. Остават 30 кубика. cm, 50 cc виж си отиде.
Според осмотичните закони не трябва да има преход. Ако премахнем жизнените свойства от чревната стена с натриев флуорид, тогава разтворът няма да изчезне.
Нека изсипем кръвен серум в същото черво. Междувременно, връщайки се към изложението, ще кажа, че тези експерименти на Хайденхайн остават доста валидни и до днес. Тези експерименти доказват, че процесът на абсорбиране е твърде сложен, за да бъде обхванат от известните физични закони. Положението на действието на тези закони е толкова сложно тук, че тези физико -химични закони са скрити за нас и явлението изглежда не е в съгласие със законите на физиката. Законите, разбира се, имат приложение тук, но те не се виждат от нас. Това показва, че там, където познаваме добре материята, съществува пълно господство на физиката и химията, а там, където знаем малко, има противоречие, което само разкрива нашето невежество и нищо друго.
Така че процесът на усвояване е сложен процес Сега ще преминем в детайлите на прехвърлянето на хранителни вещества в тялото. Как по какви пътища преминават веществата? Тук има няколко начина, но има два основни. Ще ви припомня кратка хистология на червата. Цялата лигавица на червата е осеяна с издатини, ворсинки. Те имат сложен дизайн. Вътре във всяка от ворси има централна кухина. Различни елементи са разположени на повърхността на ворсинките. Започвайки отвътре, първо има слой от цилиндричен епител, който има своеобразно разположение на външната част под формата на надлъжно набраздена граница, след това клетъчното тяло и ядрото. Този ред е последван от скелета на съединителната тъкан, основата. В тази основа точно под клетките са капилярите на кръвоносните съдове. След това серия от слотове, които провеждат течности дълбоко в централния канал. В същата основа на съединителната тъкан има нерви. Ето най -общо състава на ворсинките. Централната част на ворсинките е началото на специални тръби, така наречените млечни съдове, които бяха обсъдени по-рано, когато се говори за значението на жлъчката. Млечните съдове са началото на лимфната система. Първоначално те са много малки, така че могат да се видят само под микроскоп, а след това се превръщат в съдове с такъв размер, че да ги виждаме с просто око. Следователно, за течността, преминаваща през вилите и през цялата лигавица, има възможност да отиде на две места: или да премине през слоя от колонен епител и съединителната тъкан и да проникне в млечните съдове, или да влезе в кръвоносната система, в капилярите, които са във вилиците, лежат под слой от цилиндрични клетки. Що се отнася до материята, тогава има два начина; или в централните канали на ворсинките и следователно след това в млечните съдове, или в капилярите, в кръвта.
Сега въпросът. Какво се сервира къде? Какви преработени и абсорбирани вещества влизат в кръвта и какво в лимфата? Този въпрос може да бъде решен по този начин: трябва да вземете или кръв, или съдържанието на млечен съд - млечен сок - и да анализирате състава им, след като сте дали някакви вещества на животното за храна. Това е чисто химическа задача. Сега нека ви напомня, че изтичащата кръв, изтичащата кръв, отива от червата по специален клон, по порталната система. Порталната система се състои от вени, които събират кръв от храносмилателния канал. Те не отиват директно към сърцето, а първо отиват в черния дроб, разбиват се там в капиляри, отново се събират в големи съдове и след това се появяват в долната куха вена. Следователно, за такъв анализ трябва да се вземе кръв от порталната система. За да разберете какво е паднало в съдовете за мляко, трябва да направите това. Тези съдове в началото са много малки, трудно е да се работи с тях, трудно е да се вмъкне тръба в тях. Следователно трябва да вземете съдовете там, където те вече са достатъчно големи. Млечните съдове се сливат с лимфната система, която преминава през всички части на тялото. По този начин млечните съдове са един от клоните на лимфната система. След сливането с останалите лимфни съдове, млечните съдове продължават да се разширяват и в крайна сметка огромно количество лимфа и млечен сок се събират и текат в голям съд. Това е така нареченият гръден канал - ductus thoracicus. Тук се появява и абсорбираната течност. Тук лесно можете да го получите. Можем да отворим този гръден канал и след това да изпомпваме млечната течност от корема в него по желание.
Следователно има пълна възможност за наблюдение на абсорбираните вещества или в кръвта, или в дуктуса на гръдния кош.
Сега да видим резултатите от експеримента. Наляха се 90 кубически метра. вижте кръвен серум в йеюнума. Остават 65 кубически метра. см, следователно 25 кубически метра. вижте течността, изхвърлена от червата. Излезе течност, която по състав е абсолютно същата като течността от другата страна на червата. Защо не се получи достатъчно? Това се обяснява с факта, че колкото повече експерименти правим на това черво, толкова по -дълго отнемат тези експерименти, толкова повече червата се отдалечава от нормалните условия и толкова по -лошо работи. Освен това има и други, по -дълбоки причини, за които няма да говоря сега.
Популярни статии на сайта от раздела „Медицина и здраве“
|
.
|
Дебелото черво (intestinalinum crassum) следва тънките черва. В дебелото черво изолацията на сляпото черво, дебелото черво и ректума. Дебелото черво от своя страна е представено от възходящо дебело черво, напречно, низходящо и сигмовидно дебело черво. Функцията на дебелото черво е да абсорбира вода, да образува и отделя изпражнения - неразградени остатъци от храна. Дебелото черво е с дължина приблизително 160 см. При живите хора е малко по -дълго поради високата еластичност на тъканите. Дължината на сляпото черво при възрастен е 4,66% от цялата дължина на дебелото черво. Дължината на възходящото дебело черво е 16,17%, напречното дебело черво е 34,55%, низходящото дебело черво е 13,72%, а сигмоидното дебело черво е 29,59%от дължината на дебелото черво при възрастни (с изключение на ректума). Диаметърът на дебелото черво варира индивидуално, средно е 5-8 cm и намалява в посока от цекума към ректума. Масата на дебелото черво (без съдържание) при възрастен е приблизително 370 g.
Слепото черво (цекум) е началната част на дебелото черво, илеумът се изпразва в него. Слепото черво има сакулирана форма, свободен купол, обърнат надолу, от който надолу се простира червеобразен апендикс (апендикс).
По -рядко сляпото черво е конусовидно. Дължината на сляпото черво е 4-8 см. Задната повърхност на слепото черво е разположена върху илеума и псоас големите мускули. Предната повърхност на червата е в непосредствена близост до предната коремна стена. Слепото черво няма мезентерия, но перитонеума е покрит от всички страни (интраперитонеално положение). Апендиксът, който е важен орган на имунната система, е анатомично и топографски свързан с цекума.
Възходящото дебело черво (colon ascendens) има дължина 18-20 см. Положението на възходящото дебело черво е променливо. Задната му стена заема крайно дясното странично положение на задната стена на коремната кухина. Червата са насочени вертикално нагоре, разположени първо отпред на квадратния мускул на долната част на гърба, след това отпред от десния бъбрек, лежащ ретроперитонеално. Близо до долната (висцерална) повърхност на черния дроб възходящото дебело черво образува завой наляво и напред и преминава в напречното дебело черво. Това е дясното (чернодробно) огъване на дебелото черво (flexura coli dextra).
Напречното дебело черво (colon transversum) обикновено провисва надолу по дъгообразен начин. Началото му е в областта на дясното подребрие (дясно чернодробно огъване) на нивото на Х реберния хрущял, след което червата отива в коса посока отдясно наляво, първо надолу, след това нагоре в областта на лявото подребрие. Дължината на напречното дебело черво е приблизително 50 cm (25 до 62 cm).
Спускащото се дебело черво (colon descendens) започва от левия завой на дебелото черво надолу и преминава в сигмоидното дебело черво на нивото на илиачния гребен на илиума. Дължината на низходящото дебело черво е средно 23 см (10 до 30 см). Спускащото се дебело черво се намира в лявата част на корема.
Сигмоидното дебело черво (colon sigmoideum) започва на нивото на левия илиачен гребен и преминава в ректума на нивото на носа на сакрума. Дължината на червата варира от 15 до 67 см (средно - 54 см). Сигмовидното дебело черво образува 1-2 бримки (завои), които лежат пред крилото на лявата илиачна кост и частично се спускат в тазовата кухина. Сигмоидното дебело черво се намира интраперитонеално, има мезентерия. Наличието на мезентерията причинява значителна подвижност на сигмоидното дебело черво.
Характерна външна особеност на цекума и дебелото черво е наличието на три мускулни ленти - ленти на дебелото черво (taeniae coli), всяка с ширина 3-6 мм. Свободните, мезентериални и саментни ленти започват в основата на апендикса и отиват в началото на ректума. Лентите се образуват поради концентрацията на надлъжния мускулен слой в три участъка от стената на дебелото черво (в областта на лентите).
- Мезентериалната лента (taenia mesocolica) съответства на мястото на прикрепване към дебелото черво (към напречното дебело черво и сигмовидното дебело черво) на тяхната мезентерия или линията на прикрепване на червата (възходящо дебело черво и низходящо) към задната коремна стена.
- Елементарната лента (taenia omentalis) е разположена на предната повърхност на напречното дебело черво, където към него е прикрепен по -големия оментум, и на местата на ементални процеси в други части на дебелото черво.
- Свободната лента (taenia libera) е разположена на предните (свободни) повърхности на възходящото дебело черво и низходящото дебело черво и на долната повърхност на напречното дебело черво поради отпускането му и лекото усукване около надлъжната ос.
Стените на дебелото черво се характеризират с наличието на ушни процеси-пръстовидни, изпълнени с мазнини издатини, покрити с висцерален перитонеум. Дължината на процесите е 3-5 см, а броят им се увеличава в дисталната посока. Епиплоичните процеси (appenditions epiploicae) играят амортизираща роля (вероятно) по време на перисталтиката (стойност на буфера), служат като депа за мазнини в тялото. По цялото дебело черво, поради по -късата дължина на мускулните ленти в сравнение със стените на съседни части на органа, в червата се образуват издатини - haustra coli.
Стената на дебелото черво се състои от лигавицата, субмукозата, мускулната и серозната (адвентиция) мембрана.
Лигавицата на дебелото черво (tunica mucosa) се характеризира със значителен брой напречни полумесечни гънки. Височината на лунните гънки (plicae semilunares) варира от няколко милиметра до 1-2 см. Гънките се образуват от лигавицата и субмукозата в зоните между панделките на червата. Ректумът, в горния си участък (ампула), също има напречни гънки (plicae transversae recti). В долната част (анален канал) има 8-10 надлъжни гънки. Това са аналните (анални) колони (columnnae anales). Между аналните стълбове има канали - аналните (анални) синуси или синусите (синусовите анали). По стените на тези синуси се отварят отделителните канали на 5-38 многоклетъчни алвеоларно-тубуларни лигавични анални жлези, чиито основни участъци са разположени в субмукозата на аналния канал. Линията, на нивото на която са свързани долните краища на аналните колони и синусите със същото име, се нарича ректално-анална линия (hnea anorectalis).
Лигавицата на дебелото черво е облицована с еднослоен призматичен епител. Той е представен от три типа клетки: колонови епителни клетки (абсорбционни клетки), бокаловидни екзокриноцити и ендокриноцити. На нивото на аналния канал монослойният епител се заменя със стратифициран кубичен епител. По-дистално има рязък преход от многослоен кубичен към многослоен плосък некератинизиращ и постепенно към кератинизиращ епител.
Собствената ламина на лигавицата на дебелото черво се образува от рохкава влакнеста съединителна тъкан. В дебелината му има 7,5-12 милиона жлези на дебелото черво (криптите на Либеркун), които изпълняват не само секреторна, но и абсорбираща функция. В стените на сляпото черво се намират 4,5% от жлезите, в стените на дебелото черво - 90% и в ректума - 5,5% от жлезите. Разпределението на жлезите на дебелото черво има свои собствени характеристики. Плътността на местоположението им на нивото на лентите на дебелото черво е по-висока (с 4-12%), отколкото между лентите. Размерът на жлезите се увеличава на върха на полулунните гънки, както и в зоните на сфинктера на червата (в сравнение с междусфинктеричните зони). Стените на жлезите са представени от еднослоен епител, разположен върху базалната мембрана. Сред епителните клетки на жлезите преобладават бокални и абсорбционни клетки. Постоянно се срещат недиференцирани (стволови) и непостоянни ендокринни клетки. Броят на ендокриноцитите се увеличава в посока от цекума към ректума. Сред тях са EC клетки (образуват серотонин и мелатонин), D 2 клетки (секретират вазоинтестинален полипептид), А клетки (секретират глюкагон).
По протежение на lamina propria на лигавицата на дебелото черво има 5,5-6 хиляди единични лимфоидни възли, лимфоидни и мастоцити, а понякога и няколко еозинофили и неутрофили. Единични лимфоцити присъстват и в епителната лигавица на червата. В дебелината на lamina propria на лигавицата има кръвни и лимфни капиляри и съдове, немиелинизирани нервни клетки на интрамуралния нервен сплит и нервни влакна.
Мускулната плоча на лигавицата е представена от снопове гладкомускулни клетки, образуващи два слоя. Вътрешният слой е ориентиран кръгово, външният - косо и надлъжно. От мускулната плоча в дебелината на lamina propria на лигавицата се отделят снопове гладкомускулни клетки с дължина 10-30 µm, диаметър 0,2-2,0 µm. Тънките мускулни снопове обграждат жлезите на дебелото черво и допринасят за елиминирането на техните секрети.
Подмукозата (tela submucosa) се образува от рохкава влакнеста съединителна тъкан, в дебелината на която са разположени лимфоидни възли, подмукозен нервен (Мейснер) сплит, кръвни и лимфни капиляри, лигавични жлези (на нивото на аналния канал).
Мускулната мембрана (tunica muscularis) на дебелото черво, чиято дебелина се увеличава в посока от цекума към ректума, се състои от два мускулни слоя - кръгови (вътрешни) непрекъснати и надлъжни (външни) - под формата на три ленти на сляпото черво и дебелото черво. Между тези слоеве е междумускулният нервен (Auerbach) сплит, представен от ганглиозни клетки, глиоцити (Schwann и сателитни клетки) и нервни влакна. Ганглиозните клетки преобладават количествено в зоните, съответстващи на лентите на дебелото черво. Вътрешната част на кръговия слой е зоната на образуване на перисталтични вълни, които се генерират от интерстициални нервни клетки на Cajal, разположени в дебелината на субмукозата на границата с гладките мускули на дебелото черво.
На някои места, особено в зоната на преход от един участък на дебелото черво към друг, има слабо изразено удебеляване на кръгово ориентирани снопове гладка мускулатура. На тези места в процеса на храносмилане се наблюдава стесняване на чревния лумен, които се наричат функционални сфинктери на дебелото черво, които регулират преминаването на чревното съдържание. Разпределете цецино-възходящия сфинктер, разположен на нивото на горния ръб на илеално-цекологичната клапа. Следващият сфинктер на Хирш образува стесняване на дебелото черво в областта на десния му завой (чернодробно). По дължината на напречното дебело черво са дефинирани три функционални сфинктера, а десният сфинктер е разположен в началната част на напречното дебело черво. Средният напречен количен сфинктер и сфинктерът на лявото оръдие са разположени по -близо до лявата (далака) огъване на дебелото черво. Директно в областта на левия завой на дебелото черво е Pyra сфинктер. С прехода на низходящото дебело черво към сигмоидното дебело черво има низходящ сигмоиден сфинктер. В рамките на сигмоидното дебело черво горният и долният сигмоиден сфинктер са изолирани. Сигмоидно-ректалният сфинктер (О "Берние) се намира на границата на тези две части на дебелото черво.
Туниката сероза обхваща дебелото черво по различни начини. Сляпото, напречното дебело черво, сигмоидната и горната част на ректума са покрити от перитонеума от всички страни. Тези участъци на дебелото черво са разположени интраперитонеално (интраперитонеално). Възходящото дебело черво и низходящото дебело черво, както и средната част на ректума, са частично покрити от перитонеума от три страни (мезоперитонеално). Долната част на ректума не е покрита от перитонеума. Външната обвивка на тази част на червата е адвентиция. Перитонеумът (tunica serosa), обхващащ дебелото черво, при преминаване към стените на коремната кухина или към съседни органи образува мезентерия, множество гънки (така наречените връзки на дебелото черво). Тези гънки (връзки) изпълняват функциите на фиксиращ апарат, предотвратяват изместването и пролапса на червата, служат като начини за допълнително кръвоснабдяване на червата през кръвоносните съдове, преминаващи през тях. Броят на такива връзки варира индивидуално. Горната илеално-цекална гънка (plica iliocaecalis superior) е продължение на мезентерията на тънките черва вдясно. Той се прикрепя към медиалната повърхност на началната част на възходящото дебело черво, а основата му е свързана с перитонеума на десния мезентериален синус. на триъгълна формация до десния ръб на стената на входа на малкия таз. При жените лигаментът преминава към поддържащия лигамент на яйчника; при мъжете той отива в дълбокия пръстен на ингвиналния канал, където постепенно преминава в париеталния (париетален) перитонеум. Левият френо-дебелочревен лигамент (lig.phrenocolicum sinistrum) се намира между реберната част на диафрагмата и левия завой на дебелото черво. В долната част лигаментът се простира до областта на ъгъла на далака, образуван от напречното дебело черво и низходящото дебело черво, свързвайки ги един с друг. Обикновено този лигамент е снаден с голям омент. Останалите връзки са непостоянни. Те по -често фиксират зоните на преход от един участък на дебелото черво към друг.
Вие също ще се интересувате от:
Бронхопневмония: клинична и рентгенова картина Клинична картина. Започнете ...

Блокерите на бавните калциеви канали (BMCC) са група лекарства, които ...

Всички материали на сайта са подготвени от специалисти в областта на хирургията, анатомията и специализирани ...

Костите непрекъснато се обновяват. Старите структури се разрушават от остеокласти и ...
Разликата между традиционните и селективните (селективни) болкоуспокояващи е почти винаги ...

