



Приблизително 1 на 150 деца се ражда с хромозомна аномалия. Тези нарушения са причинени от грешки в броя или структурата на хромозомите. Много деца с хромозомни проблеми имат умствени и/или физически вродени дефекти. Някои хромозомни проблеми в крайна сметка водят до спонтанен аборт или мъртво раждане.
Хромозомите са нишковидни структури, намиращи се в клетките на нашето тяло и съдържащи набор от гени. Хората имат около 20–25 хиляди гена, които определят характеристики като цвета на очите и косата и също така са отговорни за растежа и развитието на всяка част от тялото. Всеки човек обикновено има 46 хромозоми, събрани в 23 хромозомни двойки, в които една хромозома се наследява от майката, а втората се наследява от бащата.
Причини за хромозомни аномалии
Хромозомните аномалии обикновено са резултат от грешка, възникнала по време на узряването на спермата или яйцеклетката. Защо възникват тези грешки все още не е известно.
Яйцата и спермата обикновено съдържат 23 хромозоми. Когато се съберат, те образуват оплодена яйцеклетка с 46 хромозоми. Но понякога нещо се обърка по време на (или преди) оплождането. Например, яйцеклетката или спермата може да се развият неправилно, в резултат на което те могат да имат допълнителни хромозоми или, обратно, да им липсват хромозоми.
В този случай клетки с грешен брой хромозоми са прикрепени към нормална яйцеклетка или сперма, в резултат на което полученият ембрион има хромозомни аномалии.
Най-често срещаният тип хромозомна аномалиянаречена тризомия. Това означава, че вместо да има две копия на определена хромозома, човек има три копия. Например, те имат три копия на хромозома 21.
В повечето случаи ембрион с грешен брой хромозоми не оцелява. В такива случаи жената има спонтанен аборт, обикновено в ранните етапи. Това често се случва много рано по време на бременност, преди жената дори да разбере, че е бременна. Повече от 50% от спонтанните аборти през първия триместър са причинени от хромозомни аномалии в ембриона.
Други грешки могат да възникнат преди оплождането. Те могат да доведат до промени в структурата на една или повече хромозоми. Хората със структурни хромозомни аномалии обикновено имат нормален брой хромозоми. Въпреки това, малки части от хромозома (или цяла хромозома) могат да бъдат изтрити, копирани, обърнати, неправилно поставени или заменени с част от друга хромозома. Тези структурни пренареждания може да нямат ефект върху човек, ако той има всички хромозоми, но те просто се пренареждат. В други случаи такива пренареждания могат да доведат до загуба на бременност или вродени дефекти.
Грешки в клетъчното делене могат да възникнат скоро след оплождането. Това може да доведе до мозаицизъм, състояние, при което човек има клетки с различен генетичен състав. Например, хората с една форма на мозаицизъм, синдром на Търнър, нямат X хромозома в някои, но не във всички клетки.
Диагностика на хромозомни аномалии
Хромозомните аномалии могат да бъдат диагностицирани преди раждането на бебето чрез пренатално изследване, като амниоцентеза или хорионбиопсия, или след раждането чрез кръвен тест.
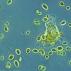
Клетките, получени от тези тестове, се отглеждат в лаборатория и след това техните хромозоми се изследват под микроскоп. Лабораторията прави изображение (кариотип) на всички хромозоми на човек, подредени в ред от най-големите към най-малките. Кариотипът показва броя, размера и формата на хромозомите и помага на лекарите да идентифицират всякакви аномалии.
Първият пренатален скрининг се състои в вземане на кръвен тест на майката през първия триместър на бременността (между 10 и 13 седмица от бременността), както и специално ултразвуково изследване на тила на бебето (т.нар. нухална транслуценция).
Вторият пренатален скрининг се извършва през втория триместър на бременността и се състои от кръвен тест на майката между 16 и 18 седмици. Този скрининг идентифицира бременности, които са изложени на по-висок риск от генетични заболявания.
Скрининговите тестове обаче не могат точно да диагностицират синдрома на Даун или други. Лекарите предлагат жените, които имат необичайни резултати от скринингови тестове, да се подложат на допълнителни тестове - хорионбиопсия и амниоцентеза - за окончателно диагностициране или изключване на тези нарушения.
Най-честите хромозомни аномалии
Първите 22 двойки хромозоми се наричат автозоми или соматични (неполови) хромозоми. Най-честите аномалии на тези хромозоми включват:
1. Синдром на Даун (тризомия 21) е една от най-честите хромозомни аномалии, диагностицирана при приблизително 1 на 800 бебета. Хората със синдром на Даун имат различна степен на умствено развитие, характерни черти на лицето и често вродени аномалии в развитието на сърцето и други проблеми.
Съвременните перспективи за развитие на деца със синдром на Даун са много по-ярки, отколкото преди. Повечето от тях са с леки до умерени интелектуални затруднения. С ранна интервенция и специално образование много от тези деца се научават да четат и пишат и участват в различни дейности от детството нататък.
Рискът от синдром на Даун и други тризомии се увеличава с възрастта на майката. Рискът от раждане на дете със синдром на Даун е приблизително:
- 1 на 1300 – ако майката е на 25 години;
- 1 на 1000 – ако майката е на 30 години;
- 1 на 400 – ако майката е на 35 години;
- 1 на 100 – ако майката е на 40 години;
- 1 на 35 – ако майката е на 45 години.
2. Тризомия 13 и 18 хромозома – тези тризомии обикновено са по-сериозни от синдрома на Даун, но за щастие са доста редки. Около 1 на 16 000 бебета се раждат с тризомия 13 (синдром на Патау), а 1 на 5000 бебета се раждат с тризомия 18 (синдром на Едуардс). Децата с тризомия 13 и 18 обикновено страдат от тежка умствена изостаналост и много вродени дефекти. Повечето от тези деца умират преди да навършат една година.
Последната, 23-та двойка хромозоми са половите хромозоми, наречени X хромозоми и Y хромозоми. Обикновено жените имат две X хромозоми, докато мъжете имат една X хромозома и една Y хромозома. Аномалиите на половите хромозоми могат да причинят безплодие, проблеми с растежа и проблеми с ученето и поведението.
Най-честите аномалии на половите хромозоми включват:
1. Синдром на Търнър – Това заболяване засяга приблизително 1 на 2500 женски фетуса. Момиче със синдром на Търнър има една нормална Х хромозома и напълно или частично липсва втора Х хромозома. Обикновено тези момичета са безплодни и няма да претърпят промените на нормалния пубертет, освен ако не приемат синтетични полови хормони.
Момичетата, засегнати от синдрома на Търнър, са много ниски, въпреки че лечението с хормон на растежа може да помогне за увеличаване на височината. Освен това те имат цял набор от здравословни проблеми, особено със сърцето и бъбреците. Повечето момичета със синдром на Търнър имат нормална интелигентност, въпреки че изпитват някои затруднения в ученето, особено по математика и пространствено мислене.
2. Тризомия X хромозома – Около 1 на 1000 жени имат допълнителна X хромозома. Такива жени са много високи. Те обикновено нямат физически вродени дефекти, имат нормален пубертет и са плодовити. Такива жени имат нормален интелект, но може да имат и сериозни проблеми с ученето.
Тъй като такива момичета са здрави и имат нормален външен вид, родителите им често не знаят, че дъщеря им го има. Някои родители разбират, че детето им има подобно заболяване, ако майката е била подложена на един от инвазивните пренатални диагностични методи (амниоцентеза или хориоцентеза) по време на бременност.
3. Синдром на Клайнфелтер – Това разстройство засяга приблизително 1 на 500 до 1000 момчета. Момчетата със синдром на Клайнфелтер имат две (а понякога и повече) X хромозоми заедно с една нормална Y хромозома. Такива момчета обикновено имат нормален интелект, въпреки че много от тях имат проблеми с ученето. Когато такива момчета пораснат, те имат намалена секреция на тестостерон и са безплодни.
4. Дисомия на Y хромозомата (XYY) – Около 1 на 1000 мъже се раждат с една или повече допълнителни Y хромозоми. Тези мъже имат нормален пубертет и не са безплодни. Повечето имат нормален интелект, въпреки че може да има някои затруднения в обучението, поведенчески затруднения и проблеми с речта и усвояването на езика. Както при тризомията X при жените, много мъже и техните родители не знаят, че имат разстройството до пренатална диагностика.
По-рядко срещани хромозомни аномалии
Новите методи за анализ на хромозомите могат да открият малки хромозомни аномалии, които не могат да се видят дори под мощен микроскоп. В резултат на това все повече родители научават, че детето им има генетична аномалия.
Някои от тези необичайни и редки аномалии включват:
- Делеция - липса на малък участък от хромозома;
- Микроделеция - липсата на много малък брой хромозоми, може би липсва само един ген;
- Транслокация - част от една хромозома се присъединява към друга хромозома;
- Инверсия - част от хромозомата се пропуска и редът на гените се обръща;
- Дублиране (удвояване) - част от хромозомата се дублира, което води до образуването на допълнителен генетичен материал;
- Пръстенова хромозома – когато генетичният материал се отстрани от двата края на хромозомата и новите краища се съединят, за да образуват пръстен.
Някои хромозомни патологии са толкова редки, че само един или няколко случая са известни на науката. Някои аномалии (например някои транслокации и инверсии) може да нямат ефект върху здравето на човека, ако липсва негенетичен материал.
Някои необичайни нарушения могат да бъдат причинени от малки хромозомни делеции. Примери за това са:
- Синдром на Cry Cat (изтриване на хромозома 5) - болните деца в ранна детска възраст се отличават с висок вик, сякаш котка крещи. Те имат значителни проблеми във физическото и интелектуалното развитие. Приблизително 1 на 20-50 хиляди бебета се раждат с това заболяване;
- Синдром на Прадер-УилИ (изтриване на хромозома 15) - болните деца имат отклонения в умственото развитие и ученето, нисък ръст и поведенчески проблеми. Повечето от тези деца развиват екстремно затлъстяване. Приблизително 1 на 10-25 хиляди бебета се раждат с това заболяване;
- Синдром на DiGeorge (делеция на хромозома 22 или делеция 22q11) – Около 1 на 4000 бебета се раждат с делеция в специфична част от хромозома 22. Това заличаване причинява различни проблеми, които могат да включват сърдечни дефекти, цепнатина на устната/небцето (цепнато небце и цепнатина на устната), нарушения на имунната система, необичайни черти на лицето и проблеми с ученето;
- Синдром на Wolf-Hirschhorn (делеция на хромозома 4) – това заболяване се характеризира с умствена изостаналост, сърдечни дефекти, слаб мускулен тонус, гърчове и други проблеми. Това състояние засяга приблизително 1 на 50 000 бебета.
С изключение на хората със синдром на DiGeorge, хората с горните синдроми са безплодни. Що се отнася до хората със синдром на DiGeorge, тази патология се наследява с 50% при всяка бременност.
Новите методи за анализ на хромозомите понякога могат да посочат къде липсва генетичен материал или къде присъства допълнителен ген. Ако лекарят знае точно къде е виновникът хромозомна аномалия, той може да оцени пълната степен на влиянието му върху детето и да даде приблизителна прогноза за развитието на това дете в бъдеще. Често това помага на родителите да решат да продължат бременността и да се подготвят предварително за раждането на бебе, което е малко по-различно от всички останали.
Хромозомните мутации (наричани още аберации, пренареждания) са непредвидими промени в структурата на хромозомите. Най-често се причиняват от проблеми, възникващи по време на клетъчното делене. Излагането на иницииращи фактори на околната среда е друга възможна причина за хромозомни мутации. Нека да разберем какви прояви могат да бъдат този вид промени в структурата на хромозомите и какви последствия имат за клетката и целия организъм.
Мутации. Общи положения
В биологията мутацията се определя като трайна промяна в структурата на генетичния материал. Какво означава "постоянен"? Наследява се от потомци на организъм, който има мутирала ДНК. Това става по следния начин. Една клетка получава грешна ДНК. Той се дели и две дъщери копират напълно структурата му, тоест съдържат и променен генетичен материал. След това има все повече и повече такива клетки и ако организмът премине към възпроизвеждане, неговите потомци получават подобен мутантен генотип.
Мутациите обикновено не преминават без следа. Някои от тях променят тялото толкова много, че резултатът от тези промени е смърт. Някои от тях принуждават тялото да функционира по нов начин, намалявайки способността му да се адаптира и водейки до сериозни патологии. И много малък брой мутации са от полза за тялото, като по този начин увеличават способността му да се адаптира към условията на околната среда.
Мутациите се делят на генни, хромозомни и геномни. Тази класификация се основава на различията, възникващи в различните структури на генетичния материал. По този начин хромозомните мутации засягат структурата на хромозомите, генните мутации засягат последователността на нуклеотидите в гените, а геномните мутации правят промени в генома на целия организъм, добавяйки или изваждайки цял набор от хромозоми.
Нека поговорим за хромозомните мутации по-подробно.
Какви видове хромозомни пренареждания могат да възникнат?
В зависимост от това как са локализирани промените, се разграничават следните видове хромозомни мутации.
- Интрахромозомно - трансформация на генетичен материал в рамките на една хромозома.
- Интерхромозомни - пренареждания, в резултат на които две нехомоложни хромозоми обменят своите участъци. Нехомоложните хромозоми съдържат различни гени и не се появяват по време на мейозата.
Всеки от тези видове аберации съответства на определени видове хромозомни мутации.

Изтривания
Делецията е отделяне или загуба на която и да е част от хромозома. Лесно е да се досетите, че този тип мутация е интрахромозомна.
Ако най-външната част на хромозомата е отделена, делецията се нарича терминална. Ако генетичният материал се загуби по-близо до центъра на хромозомата, такова изтриване се нарича интерстициално.
Този тип мутация може да повлияе на жизнеспособността на организма. Например, загубата на част от хромозома, кодираща определен ген, осигурява на човек имунитет срещу вируса на имунната недостатъчност. Тази адаптивна мутация е възникнала преди около 2000 години и някои хора със СПИН са успели да оцелеят само защото са имали късмета да имат хромозоми с променена структура.
Дублиране
Друг вид интрахромозомна мутация е дупликацията. Това е копиране на участък от хромозома, което се получава в резултат на грешка по време на т. нар. кръстосване или кръстосване по време на клетъчното делене.
Копиран по този начин участък може да запази позицията си, да се завърти на 180° или дори да се повтори няколко пъти и тогава такава мутация се нарича амплификация.
В растенията количеството генетичен материал може да се увеличи именно чрез повтарящи се дублации. В този случай обикновено се променя способността на цял вид да се адаптира, което означава, че такива мутации са от голямо еволюционно значение.
Инверсии
Също така се отнася до интрахромозомни мутации. Инверсията е завъртане на определен участък от хромозома на 180°.

Частта от хромозомата, обърната в резултат на инверсия, може да бъде от едната страна на центромера (парацентрична инверсия) или от противоположните й страни (перицентрична). Центромерът е така наречената област на първичната констрикция на хромозомата.
Обикновено инверсиите не засягат външните признаци на тялото и не водят до патологии. Съществува обаче предположение, че при жени с инверсия на определена част от деветата хромозома вероятността от спонтанен аборт по време на бременност се увеличава с 30%.
Транслокации
Транслокацията е преместването на част от една хромозома към друга. Тези мутации са от междухромозомен тип. Има два вида транслокации.
- Реципрочен е обменът на две хромозоми в определени области.
- Robertsonian - сливане на две хромозоми с късо рамо (акроцентрично). По време на Робъртсъновата транслокация късите участъци от двете хромозоми се губят.
Реципрочните транслокации водят до проблеми с раждаемостта при хората. Понякога такива мутации причиняват спонтанен аборт или водят до раждането на деца с вродени патологии на развитието.
Робъртсъновите транслокации са доста чести при хората. По-специално, ако възникне транслокация, включваща хромозома 21, плодът развива синдром на Даун, една от най-често съобщаваните вродени патологии.
Изохромозоми
Изохромозомите са хромозоми, които са загубили едното си рамо, но са го заменили с точно копие на другото си рамо. Тоест по същество такъв процес може да се счита за изтриване и обръщане в една бутилка. В много редки случаи такива хромозоми имат две центромери.
Изохромозомите присъстват в генотипа на жените, страдащи от синдрома на Шерешевски-Търнър.
Всички видове хромозомни мутации, описани по-горе, са присъщи на различни живи организми, включително хора. Как се проявяват?
Хромозомни мутации. Примери
Мутации могат да възникнат в половите хромозоми и в автозомите (всички други сдвоени хромозоми на клетката). Ако мутагенезата засяга половите хромозоми, последствията за тялото обикновено са тежки. Възникват вродени патологии, които засягат умственото развитие на индивида и обикновено се изразяват в промени във фенотипа. Тоест, външно мутантните организми се различават от нормалните.
Геномните и хромозомните мутации се срещат по-често в растенията. Те обаче се срещат както при животни, така и при хора. Хромозомните мутации, примери за които ще разгледаме по-долу, се проявяват в появата на тежки наследствени патологии. Това са синдром на Волф-Хиршхорн, синдром на "плачещата котка", частична тризомия на късото рамо на хромозома 9, както и някои други.
Синдром на плача на котката
Това заболяване е открито през 1963 г. Възниква поради частична монозомия на късото рамо на хромозома 5, причинена от делеция. Едно на 45 000 деца се ражда с този синдром.

Защо тази болест получи такова име? Децата, страдащи от това заболяване, имат характерен вик, който наподобява мяукане на котка.
Когато късото рамо на петата хромозома бъде изтрито, различни части от нея могат да бъдат загубени. Клиничните прояви на заболяването пряко зависят от това кои гени са били загубени по време на тази мутация.
Структурата на ларинкса се променя при всички пациенти, което означава, че "котешкият вик" е характерен за всички без изключение. Повечето хора, страдащи от този синдром, изпитват промяна в структурата на черепа: намаляване на мозъчната област, лице с форма на луна. В случай на синдром на "Cry the Cat", ушите обикновено са разположени ниско. Понякога пациентите имат вродени патологии на сърцето или други органи. Умствената изостаналост също става характерна черта.
Обикновено пациентите с този синдром умират в ранна детска възраст, само 10% от тях оцеляват до десетгодишна възраст. Има обаче и случаи на дълголетие със синдрома на "вика на котката" - до 50 години.
Синдром на Wolf-Hirschhorn
Този синдром е много по-рядко срещан - 1 случай на 100 000 раждания. Причинява се от делецията на един от сегментите на късото рамо на четвъртата хромозома.

Проявите на това заболяване са разнообразни: забавено развитие на физическата и умствената сфера, микроцефалия, характерен клюновиден нос, страбизъм, цепнато небце или горна устна, малка уста, дефекти на вътрешните органи.
Подобно на много други човешки хромозомни мутации, болестта на Wolf-Hirschhorn се класифицира като полулетална. Това означава, че жизнеспособността на тялото с такова заболяване е значително намалена. Децата, диагностицирани със синдром на Wolf-Hirschhorn, обикновено не живеят след 1 година, но е регистриран един случай, при който пациентът е живял 26 години.
Синдром на частична тризомия на късото рамо на хромозома 9
Това заболяване възниква поради небалансирани дупликации в деветата хромозома, в резултат на което върху тази хромозома има повече генетичен материал. Общо са известни повече от 200 случая на такива мутации при хора.
Клиничната картина се характеризира със забавено физическо развитие, лека умствена изостаналост и характерна мимика. Сърдечни дефекти се откриват при една четвърт от всички пациенти.
При синдром на частична тризомия на късото рамо на хромозома 9 прогнозата все още е сравнително благоприятна: повечето пациенти оцеляват до старост.

Други синдроми
Понякога хромозомни мутации възникват дори в много малки участъци от ДНК. Заболяванията в такива случаи обикновено се причиняват от дупликации или делеции и се наричат съответно микродупликации или микроделеции.
Най-често срещаният такъв синдром е болестта на Прадер-Уили. Това се случва поради микроделеция на част от хромозома 15. Интересното е, че тази хромозома трябва да бъде получена от тялото от бащата. В резултат на микроделеции са засегнати 12 гена. Пациентите с този синдром имат умствена изостаналост, затлъстяване и обикновено имат малки крака и ръце.
Друг пример за такива хромозомни заболявания е синдромът на Сотос. На дългата ръка на хромозома 5 се появява микроделеция. Клиничната картина на това наследствено заболяване се характеризира с бърз растеж, увеличаване на размера на ръцете и краката, наличие на изпъкнало чело и известно умствено изоставане. Честотата на този синдром не е установена.
Хромозомни мутации, по-точно микроделеции в области на хромозоми 13 и 15, причиняват съответно тумор на Wilms и ретинбластом. Туморът на Wilms е рак на бъбрека, който се среща предимно при деца. Ретинобластомът е злокачествен тумор на ретината, който се среща и при деца. Тези заболявания са лечими, ако се диагностицират в ранните етапи. В някои случаи лекарите прибягват до хирургическа намеса.

Съвременната медицина елиминира много заболявания, но все още не е възможно да се излекуват или поне да се предотвратят хромозомни мутации. Те могат да бъдат открити само в началото на развитието на плода. Генното инженерство обаче не стои неподвижно. Може би скоро ще бъде намерен начин за предотвратяване на заболявания, причинени от хромозомни мутации.
Различните клетки на един и същи организъм и различни индивиди от един и същи вид имат като правило еднакъв брой хромозоми, с изключение на гаметите, които имат наполовина по-малко хромозоми от соматичните клетки. В допълнение, броят на хомоложните гени и редът на гените в тях също, като правило, съвпадат в различни клетки и в различни представители на един и същи вид. Въпреки това броят на хромозомите, техният размер и организация варират значително при различните видове. Хаплоидният геном на повечето животни съдържа около 2109 bp. (нуклеотидни двойки); при някои насекоми и примитивни хордови това число е само 108, докато при някои земноводни, напротив, достига 1011 bp. на ядро. Количеството ДНК в растителните клетки варира в още по-широки граници. ДНК е част от хромозомите, чийто брой може да варира значително: в клетките на нематоди Parascarisunivalensсъдържа една двойка хромозоми, докато пеперудата Лизандра атлантскаброят на хромозомите е приблизително 220, а при папратите Ophioglossum reticulayumнадхвърля 600.
По време на еволюцията на организма не само броят и размерът на хромозомите могат да се променят, но и тяхната организация: отделни участъци от хромозоми могат да променят местоположението си в хромозомата и дори да се преместват от една хромозома в друга. Промените в броя, размера и организацията на хромозомите се наричат хромозомни мутации, пренарежданияили аберации[Аяла]. Те представляват движения на генетичен материал, водещи до промени в структурата на хромозомите в кариотипа. Такива пренареждания могат да включват участъци от една хромозома или различни - нехомоложни - хромозоми. В съответствие с този критерий се разграничават интрахромозомни и междухромозомни аберации.
Хромозомните пренареждания често водят до различни фенотипни промени, които се обясняват с локализирането на точки на прекъсване в или близо до определени гени.
Класификация на хромозомните мутации:
А. Промени в хромозомната структура.Такива промени могат да повлияят брой генив хромозомите (делеции и дупликации) и локализациягени в хромозомите (инверсии и транслокации).
1. Изтриване, или недостиг. Част от хромозома се губи.
2. Дублиране, или удвояване. Една от хромозомните области е представена повече от веднъж в хромозомния набор.
3. Инверсия. В един от регионите на хромозомата гените са разположени в обратна последователност в сравнение с нормалната. Обърнатият регион на хромозомата може или не може да включва центромер; в първия случай инверсията се нарича перицентрична (т.е. обхващаща центромера), а във втория - парацентрична (т.е. "близо до центромера").
4. Транслокация. Позицията на която и да е част от хромозомата в хромозомния набор е променена. Най-често срещаният тип транслокации са реципрочни, при които регионите се обменят между две нехомоложни хромозоми. Част от хромозома може също да промени позицията си без реципрочен обмен, като остане в същата хромозома или бъде включена в друга. Транслокации от този тип понякога се наричат транспозиции.
Б. Промени в броя на хромозомите.При промени от този вид в някои случаи (сливания и разкъсвания) общото количество на наследствения материал остава непроменено, а в други (анеуплоидия, моноплоидия и полиплоидия) се променя.
1. Центричен синтез. Две нехомоложни хромозоми се сливат в една.
2. Центрично разделение. Една хромозома се разделя на две и трябва да се образува нова центромера, в противен случай хромозомата без центромера се губи по време на клетъчното делене.
3. Анеуплоидия. В нормален хромозомен набор една или повече хромозоми или липсват, или присъстват една или повече допълнителни хромозоми.
4. Моноплоидия и полиплоидия. Броят на наборите от нехомоложни хромозоми се различава от две [Ayala].
Изтривания и недостатъци
изтриване,или дефицит, е загубата на част от хромозома. Това беше заличаването, което беше първият пример за хромозомно пренареждане, открито през 1917 г. от Бриджис с помощта на генетичен анализ. Това заличаване фенотипно води до назъбен ръб на крилото при Drosophila, наречен мутация прорез. Доказано е, че тази мутация е свързана с пола, доминираща и смъртоносна в хомозиготно състояние. Женски хетерозиготни за прорез, имат мутантен фенотип и женските и хемизиготните мъже, хомозиготни за тази мутация, не са жизнеспособни. Алел бялов присъствието прорезвърху хомоложната хромозома се държи като доминираща. Други рецесивни гени, разположени в близост до бялона X хромозомата, също стават „доминиращи“ в присъствието прорез. Това очевидно доминиране на рецесивните гени се нарича псевдодоминиране, тъй като възниква само когато определен участък от хомоложната хромозома е загубен, което води до липса на алел, комплементарен на рецесивната мутация. Псевдодоминирането е един от начините за откриване на изтривания.
Делециите обикновено са смъртоносни при хомозиготите, което показва, че някои жизненоважни гени са били загубени. Много късите делеции може да не повлияят на жизнеспособността на хомозигота.
Терминалните недостатъци или недостатъци се установяват по същите критерии, но поради тяхното местоположение по време на конюгацията не се образува верига и една хромозома се оказва по-къса от другата. Примери за недостатъци са известни в много организми, включително хора. Тежка наследствена болест синдром на плачещата котка, кръстен на естеството на звуците, издавани от болни бебета, се определя по дефиниция в 5-та хромозома. Този синдром е придружен от умствена изостаналост. Децата с този синдром обикновено умират рано.
Когато хромозомният фрагмент се отдели, той обикновено се губи, ако не съдържа центромер. Фрагментът, съдържащ центромера, се репликира и неговите копия се разпределят нормално по време на клетъчното делене. Хромозомните фрагменти не се губят в случай на дифузен центромер. В този случай могат да възникнат две теломерни хромозоми.
Методът отваря големи възможности за идентифициране на делеции, дефицити и други хромозомни аберации. диференциално оцветяване на хромозомите. Основава се на факта, че някои багрила, като багрилото Giemsa, различно оцветяват различни области на хромозомите. Поради това хромозомите придобиват характерни напречни ивици. Този метод се използва за определяне на хромозомни пренареждания в метафазни хромозоми.
Дублиране
Дублиранев тесния смисъл на думата те са двойно повторение на една и съща хромозомна секция. Известни са случаи на многократни повторения или анимациивсяка област. Те също се наричат усилвания .
Дублирането може да възникне в рамките на една и съща хромозома или да бъде придружено от прехвърляне на копие на част от генетичен материал към друга хромозома. Дублирани региони често образуват тандем ( ABCBCDE…), т.е. разположени една след друга. Тандемното дублиране се нарича обърнато (или обърнато АВССВDE…), ако генните последователности в съседни региони са взаимно противоположни. Ако дублираният регион се намира в края на хромозомата, тогава дублирането се нарича терминал.
Дублиранията могат да имат фенотипни прояви. Най-известният пример е мутацията Барна Х хромозомата Drosophila melanogaster. Тази мутация показва непълно доминиране, намалявайки броя на очните фасети.
Понякога се откриват дублирания поради факта, че индивид, хомозиготен за рецесивен алел, не проявява рецесивен белег. Този факт се обяснява с факта, че съответният доминантен алел се съдържа в дублирана област на хромозомата. Върху цитологичните препарати хетерозиготността за дупликации води до образуването на бримки, подобни на тези, които възникват при хетерозиготите за делеции.
Много дублирания и делеции могат да бъдат резултат от счупвания на хромозоми. Причината за разкъсванията може да бъде йонизиращо лъчение, действието на определени химикали или вируси. Прекъсванията могат да бъдат предизвикани и от определени характеристики на структурата и функционирането на хромозомите. Изтривания и дублирания могат да възникнат и при неравномерно пресичане. Когато подобни ДНК последователности се появят в съседни области на хромозома, конюгирането на хомолози може да се случи неправилно. Пресичането на такива неправилно конюгирани хромозомни области води до образуването на гамети с дублиране или делеция. Именно по този начин възникват хемоглобините в резултат на неравномерно кръстосване Лепореи анти- Лепоре. За дублиране и изтривания или транслокации.
Дублирането на сравнително малки участъци от ДНК, състояние на няколко нуклеотида, които са част от един ген или съседни гени, се случва много често в процеса на еволюция.
Инверсии
Инверсията е завъртане на 180° на отделни участъци от хромозомата; в този случай нито броят на хромозомите, нито броят на гените във всяка хромозома се променят ( Аяла). Ако последователността от гени в оригиналната хромозома е обозначена като ABCDEF и BCD областта е претърпяла инверсия, тогава в новата хромозома гените ще бъдат разположени в последователността ADCBEF.
В зависимост от местоположението на краищата (границите) на пренареждането по отношение на центромера инверсииразделени на перицентрични, улавящи центромера и включващи го в обърнатата секция, и парацентричен,без да се включва центромерата в обърнатата област.
Инверсиите са широко разпространен начин за еволюционна трансформация на генетичен материал. Например, хората и шимпанзетата се различават по броя на хромозомите: хората имат 2n = 46, а шимпанзетата имат 2n = 48.
Инверсията води до промяна във връзката на гените, като тяхната линейна последователност е различна от тази на оригиналната форма. Този ефект може да бъде открит, ако инверсията в хомозиготата не е летална. Рецесивната смъртност често придружава инверсии в резултат на локализирането на точки на прекъсване в жизненоважни гени или като следствие от позиционни ефекти.
Друго важно последствие от инверсията е потискането на кросинговъра, ако инверсията е в хетерозигота. Това свойство на инверсиите се използва широко за създаване на балансирани линии, които са хетерозиготни за смъртоносни мутации и не се унищожават чрез пресичане на желаната хромозома.
При хетерозиготите се откриват характерни бримки на цитологични препарати въз основа на инверсии - резултат от конюгация на структурно променена и нормална хромозома. Ако на такъв цикъл, т.е. в обърнатия участък ще настъпи единичен кросингоувър, тогава в случай на парацентрична инверсия се появява един хроматид с две центромери, които ще го разрушат, когато се разминават в анафаза. Полученият центромерен фрагмент също ще бъде загубен. В резултат на това от четири гамети само две ще бъдат пълни. Само те са способни да произвеждат жизнеспособни зиготи по време на оплождането (фиг. 7, А). В случай на хетерозиготност за перицентрична инверсия, кросингоувърът не предотвратява нормалната дивергенция на всички хроматиди. Независимо от това, само два от четирите мейотични продукта отново ще бъдат завършени, тъй като две хроматиди носят делеции на някои гени.
В същото време двойното пресичане при инверсионни хетерозиготи може да доведе до образуването на напълно жизнеспособни гамети (фиг. 1, B).
Една хромозома може да носи не само една инверсия, но и две неприпокриващи се и две припокриващи се изцяло или частично. Хетерозиготността за такива сложни пренареждания също се идентифицира цитологично от естеството на хромозомната конюгация.
Ориз. 1. − Хромозомна конюгация и последствия от единичен (A) и двоен (B) кросинговър с хетерозиготност за перицентрична инверсия
Транслокации
Транслокациите са реципрочен обмен на участъци от нехомоложни хромозоми.Реципрочните транслокации представляват взаимен обмен на участъци между две нехомоложни хромозоми (фиг. 2). Ако изобразим генните последователности в оригиналните хромозоми като ABCDEF и GHIJKL, тогава в транслокационните хромозоми генните последователности могат да бъдат например ABCDKL и GHIJEF. При хомозиготите за тези транслокации, в сравнение с оригиналните хромозоми, естеството на свързването се променя: гени, които не са свързани в оригиналните хромозоми, стават свързани и обратно. В дадения пример KL гените се оказват свързани с ABCD гените и престават да бъдат свързани с GHIJ гените.

Ориз. 2. − Транслокации
В хетерозиготите за реципрочни транслокации, гените на двете транслокирани хромозоми се държат така, сякаш принадлежат към една и съща група на свързване, тъй като само гамети, съдържащи родителския набор от хромозоми, могат да образуват жизнеспособни зиготи. В допълнение, при хетерозиготи за транслокации в близост до точките на прекъсване на хромозомите, кръстосването почти не се случва: взаимното разположение на хромозомите под формата на кръст, необходимо за конюгиране на хомоложни области в мейозата, предотвратява конюгацията в близост до точките на прекъсване на хромозомите и това намалява честотата на преминаване в тези зони.
На цитологичните препарати на хетерозиготи за реципрочни транслокации в мейотичната профаза може да се наблюдава характерна структура - кръст. Появата му се дължи на факта, че се привличат хомоложни области, разположени в различни хромозоми.
Вместо двувалентни, т.е. двойки конюгиращи хромозоми се образуват квадриваленти, състоящи се от четири свързани хромозоми, всяка от които е частично хомоложна на другите хромозоми от групата. При диакинезата хиазмата се "плъзга" от центромерите към краищата на хромозомите и кръстът се трансформира в пръстен. Понякога хромозомите на пръстена се обръщат и образуват фигура от осем.
Хетерозиготите за транслокации са частично стерилни (имат намалена плодовитост), тъй като произвеждат дефектни гамети по време на процеса на мейоза. В растенията поленовите зърна, съдържащи дублирания или делеции, обикновено умират. При животните гамети с делеции или транслокации могат да участват в оплождането, но образуваните от тях зиготи обикновено умират. Въпреки това, ако дублираната или изгубена част от хромозомата е малка, тогава потомството може да е жизнеспособно.
Хетерозиготите за реципрочни транслокации са редки при животните, но широко разпространени при растенията. Типичен пример в това отношение са различните видове трепетликова гора – Oenoyhera. Например, в O. lamarkiana, от 14 хромозоми, 12 участват в реципрочни транслокации. Следователно при мейозата в това растение се наблюдават един двувалентен и многовалентен, включително останалите 12 хромозоми. При други видове трепетлика броят на хромозомите, образуващи многовалентни, варира, което отразява броя на реципрочните транслокации.
Подобно на инверсиите, транслокациите осигуряват изолиране на нови форми и насърчават дивергенцията в рамките на един вид. Специален вид транслокация, така наречената Робертсонова транслокация или сливане, води до промяна в броя на хромозомите. Ако две теломерни хромозоми се слеят в центромера, се образува една метацентрична хромозома. Този тип хромозомни пренареждания получи името си от изследователя U.R. Робъртсън, който разбра механизма на такова сливане.
Транспозиции
Транспонирането е движението на малки участъци от генетичен материал в една и съща хромозома или между различни хромозоми. Транспозициите се извършват с участието на специални мобилни или мигриращи генетични елементи.
Мигриращите генетични елементи са описани за първи път от B. McClintock през 1947 г. във връзка с изследването на хромозомните прекъсвания в царевицата. Идентифициран е миграционен Ds (дисоциатор) локус, където за предпочитане се появяват разкъсвания на хромозоми. Ds сам по себе си не причинява разкъсвания. Те се появяват в този локус само ако в генома присъства друг миграционен елемент - Ас (активатор). И двата елемента могат да бъдат загубени с честота от няколко процента в мейотичното потомство или да променят локализацията си по време на метотични деления. В този случай Ds се движи само в присъствието на Ac.
Въвеждането на Ds в непосредствена близост или вътре в C гена, който контролира цвета на семенния алейрон, доведе до инактивиране на C гена и по този начин хетерозиготните семена C/s/s се оказаха неоцветени. В присъствието на Ac, дисоциаторът (Ds) започва да се движи и понякога напуска локуса C. В резултат на това върху неоцветените семена се появяват цветни петна от алейрон.
Едва през 80-те години, благодарение на успеха на генното инженерство, беше възможно да се изолират и изследват Ac, Ds и някои други мигриращи елементи на царевицата. Оказа се, че Ds е дефектен изтрит вариант на Ac. Структурата на елемента Ac се оказа типична за мигриращите елементи, които по това време бяха изследвани предимно в бактериите, както и в Drosophila и дрождите Sacch.Cerevisiae.
По-късно мобилни генетични елементи са открити и в други еукариотни организми. Бяло-пурпурната (wc) мутация в Drosophila има същите свойства като вмъкването на E.coli IS1. Установено е, че причинява транспониране на белия ген в автозома. В този случай възникват спонтанни делеции на съседни гени на X хромозомата, разположени отляво и отдясно на wc, подобно на делеции, причинени от елемента IS1.
Генетичен елемент, наречен sexrealizer, е открит в мухи Megaselia scalaris. Мъжете са хемизиготни за този ген; при жените той липсва. Тази полова детерминанта се намира в края на една от хромозомите, превръщайки я в полова хромозома. С честота приблизително 0,1% се образуват сперматозоиди, в които детерминантата на пола се е преместила от оригиналната полова хромозома към друга, която се е превърнала в полова хромозома. Възможно е да се създадат линии, в които различни нехомоложни хромозоми са полови хромозоми.
Откриването на мобилни генетични елементи както в прокариотите, така и в еукариотите показва, че тяхното присъствие е общо свойство на всички организми. Възниква въпросът дали тези елементи имат полезни функции за организмите. Една от хипотезите е, че те са "егоистична ДНК", осигуряваща само собственото си възпроизвеждане без никаква съпътстваща полза за своя гостоприемник. Допълнителното бреме върху метаболизма на клетката може да е много малко и егоистичната ДНК може да продължи да съществува в такива организми поради способността й да се репликира по-бързо от останалата част от генома.
При хромозомните мутации, точно както при генните мутации, се случват пренареждания в хромозомите. Първите обаче, за разлика от вторите, засягат значителни части от хромозомите.
Хромозомните мутации могат да бъдат интрахромозомни пренареждания (структурата на една хромозома се променя), както и междухромозомни пренареждания (две хромозоми се променят). Механизмът за преструктуриране може да бъде различен. Разграничават се следните видове хромозомни мутации:
Делециаз- загуба на част от хромозома.
Определение- загуба на крайния участък.
Дублиране- удвояване на част от хромозома.
Хромозомни мутации: примери. Видове хромозомни мутации
Усилване- многократно повторение.
Вмъкване- вмъкване на хромозомна област.
Инверсия- завъртане на хромозомна секция на 180°. Перицентрична инверсия - завъртане на зоната, съдържаща центромера; парацентричен - несъдържащ центромер.
Транслокация- прехвърляне на регион от една хромозома в друга.
По-специално, реципрочната транслокация е обмен на секции между нехомоложни хромозоми; Робертсонова транслокация - свързване на две акроцентрични хромозоми, което води до образуването на една метацентрична (равнораменна) или субметацентрична.
Ако се появи дефлация в двата края на хромозомата, това може да доведе до образуването на пръстенна хромозома.
В резултат на хромозомни мутации могат да възникнат хромозоми с две центромери или да не съдържат нито една.
Хромозомите без центромери се наричат ацентрични фрагменти и обикновено се губят по време на клетъчното делене. Хромозомите с две центромери се наричат дицентрични (дицентрични). В анафаза те образуват така наречените мостове и се разрушават. Впоследствие в клетката те образуват хроматинови тела (микронуклеуси).
Ако няма добавяне или загуба на генетичен материал в резултат на хромозомна мутация, тогава такива пренареждания се наричат балансирани и обикновено не водят до никакви последствия.
В резултат на небалансирано пренареждане се добавя или губи генетичен материал и организмите могат да имат изразени аномалии.
При инверсии редът на гените в част от хромозомата е обърнат. Такава мутация обикновено не се проявява фенотипно.
По време на мейозата обаче кросингоувърът може да доведе до образуването на гамети с небалансиран генетичен материал.
Хромозомните мутации възникват както в зародишните, така и в соматичните клетки. В първия случай те най-често водят до вродени заболявания и загуба на оплодителната способност.
Хромозомните пренареждания в соматичните клетки могат да доведат до рак. Успешните за даден организъм хромозомни мутации са редки, но играят важна роля в еволюционния процес и водят до образуването на нови видове.
Хромозомните мутации възникват поради появата на двуверижни скъсвания на ДНК в клетки, които не са били поправени нормално.
Такива разкъсвания възникват както спонтанно, така и под въздействието на мутагени (например йонизиращо лъчение).
Хромозомни инверсии. Хромозомни транслокации.
Инверсии- пренареждания, чиято същност е завъртане на 180 ° на зоната, образувана в резултат на две прекъсвания, със съответната промяна в местоположението на гените.
Инверсиите могат да бъдат 1) парацентрични (не включват центромера в обърнатата област, тъй като се появяват на водното рамо на хромозомата) и 2) перицентрични (включват центромера).
Този вид реконструкциянай-често се срещат в естествени популации. Група от гени, локализирани в обърната област, се предават от поколение на поколение като единичен блок, който не се разрушава чрез кръстосване. Има особено много данни за разпространението на инверсиите в популациите на мухи, комари и мушици. Наличието на инверсии в тях се установява лесно чрез микроскопско изследване на политенните хромозоми на слюнчените жлези.
Н. П. Дубинин, Н. Н. Соколов и Г.Г.
Хромозомна мутация при хората: какво е това и какви са последствията
Тиняков в поредица от произведения от 30-40-те години. миналия век, формулира механизмите на еволюционна трансформация на генетичен материал в резултат на този широко разпространен тип хромозомни мутации.
U хетерозиготиИнверсиите върху цитологичните препарати разкриват характерни бримки - резултат от конюгация на структурно променени и нормални хромозоми.
Ако в инвертираната област се случи едно кръстосване, тогава в случай на парацентрична инверсия ще възникне един хроматид с две центромери, които ще го „счупят“, когато се разминават в анафаза. Полученият центромерен фрагмент ще бъде загубен. В резултат на това от четири хроматиди две ще бъдат аберантни. В случай на хетерозиготност за перицентрична инверсия, еднократно пресичане не предотвратява дивергенцията на всички хроматиди. Но само две от четирите ще бъдат пълни, тъй като другите две хроматиди носят делеции и дублирания на няколко гена.
При загуба или удвояване по време на пресичанехромозомните участъци са много малки, те не влияят на жизнеспособността на гаметите и зиготите, образувани по време на тяхното сливане.

Ако се случат две събития в рамките на обърнатата област пресичане, тогава пълният набор от гени се запазва без делеции и дупликации и по този начин се осигурява жизнеспособността на рекомбинантите.
Експериментално получените инверсии се използват като " шкафчета» пресичане. Нашата статия предоставя примери за използване на линии на Drosophila с потиснато кръстосване поради наличието на инверсии за отчитане на смъртоносни мутации.
Инверсиите в човешките хромозоми водят до разрушаване гаметогенеза.
Транслокации- преместване на участъци от хромозома на нова позиция в нейните граници или обмен на участъци между различни хромозоми.
Разграничете транслокации:
1) симетричен(реципрочен) - свързването на центричен фрагмент от една хромозома с ацентричен фрагмент от друга, т.е. взаимен обмен на секции между две нехомоложни хромозоми (реципрочната транслокация е това, което клиницистите често откриват в семейства, където се среща повече от една хромозомна аномалия).
В резултат на конюгиране в мейозата, транспонираните хромозоми в хетерозиготите, заедно с техните непренаредени хомолози, образуват характерна фигура на "транслокационен кръст". Плътното конюгиране близо до точките на прекъсване е възпрепятствано, което води до потискане на кросинговъра в тези области. Тъй като и четирите конюгиращи хромозоми имат хомоложни области, квадривалентите се образуват в мейотичната профаза.
От шестте възможни типа хаплоидни продукти, които възникват от три метода на хромозомна дивергенция, само два типа функционират нормално: тези, които са получили пълни набори от гени, характерни за оригиналните родителски форми. Останалите четири вида гамети ще имат небалансирани хромозомни комплекти: гаметата ще съдържа хромозома с делеция или дублиране в определени участъци;
2) асиметричен- връзки на центрични или ацентрични фрагменти, в резултат на което се образуват дицентрици, трицентрици и др.;
3) Робъртсънов- сливане на нехомоложни акроцентрични хромозоми в областта на техните центромери за образуване на една метацентрична хромозома.
Транслокациите от този тип са кръстени на У. Робъртсън, който предложи хипотезата за сливане на хромозоми, за да обясни намаляването на техния брой в хромозомния набор. Центричното сливане е често срещан тип хромозомно пренареждане при хората. Може да включва всичките пет двойки акроцентрици - хромозоми с едно дълго и второ много късо (понякога трудно за откриване) рамо.
Когато се образуват Робертсонови транслокации, заедно със загубата на къси рамена, се губят и съдържащите се в тях рибозомни РНК гени, което се потвърждава от резултатите от ДНК-РНК хибридизацията. Това обаче не е придружено от функционални аномалии и носителите на такива хромозоми са напълно здрави.
Ако по време на мейозата транслокираната хромозома влезе в зародишната клетка, зиготата ще бъде тризомична. Това е типът транслокация на синдрома на Даун, който се случва.
— Върнете се към съдържанието на раздела „Генетика."
Методи за регулиране на генната активност.
3. Неспецифична регулация на генната активност. Генна компенсация на дозата в Drosophila.
4. Компенсация на генната доза при бозайници. Съвременна теория за инактивирането на X хромозомата.
5. Регулация на генната активност на ниво репликация. Транслационна и посттранслационна регулация на генната активност.
6. Мутации.
Теоретични основи на мутационната изменчивост.
7. Геномни мутации. Хаплоидия. Полиплоидия.
8. Анеуплоидия. Нулисомия. Монозомия. Полисемия.
9. Хромозомни мутации. Изтривания. Дублиране.
10. Хромозомни инверсии. Хромозомни транслокации.
Хромозомни пренареждания или хромозомни аберациисе наричат видими промени в структурата на хромозомите. Понякога хромозомните пренареждания се наричат хромозомни мутации. Хромозомните аберации (за разлика от генните мутации) винаги са уникални и неподражаеми. Следователно, при липса на инбридинг, хромозомните аберации възникват само в хетерозиготно състояние:
- в комбинация с нормални хромозоми,
- в комбинация с други аберации.
При тясно свързано кръстосване (инбридинг) е възможно образуването на хомозиготи.
Има:
- интрахромозомни аберации (фрагментация, недостатъци, дупликации, инверсии, транспозиции),
- междухромозомни (транслокации).
Раздробяване- Това е фрагментирането на хромозомите с образуването на много различни фрагменти.
Някои организми имат полицентрични хромозоми и когато се фрагментират, всеки фрагмент получава центромер, след което може да се репликира нормално и да участва в клетъчното делене.
Край на недостига или недостатъците– загуба на крайни, теломерни области на хромозомите.
В резултат на това се образуват линейни фрагменти, които нямат центромер (линейни ацентрики). Ацентриците не участват в клетъчното делене и се губят. Липсите на вътрешни участъци или делеции са загуби на участъци от хромозоми, които не засягат теломерите. Загубените зони без центромери обикновено образуват пръстеновидни ацентрики, които също се губят.
Дублиране- Това е удвояването на хромозомните участъци.
В резултат на това възникват тандемни генни последователности, например: abcabc. Дублирането е един от начините за възникване на нови гени.
Инверсии– завъртане на хромозомните участъци на 180°.
Има:
- перицентрични инверсии (обърнатата област включва центромера),
- парацентрична (обърнатата област лежи в едно от рамената на хромозомата извън центромера).
При хетерозиготите, когато нормалните и обърнатите хромозоми се пресичат, възникват ацентрици и дицентрици; в резултат на това възникват дефектни клетки и продуктите от кросингоувъра не преминават в следващите поколения (затова инверсиите образно се наричат „кросингоувър арестери“).
По този начин инверсиите допринасят за запазването на цели блокове от гени - супергени. Ако инверсиите се комбинират с дублиране, могат да възникнат палиндроми, например: abccba.
Транспозиции- Това са движения на хромозомни участъци към други локуси (точки) на същата хромозома.
Има участъци от хромозоми, които са склонни към транспониране; те се наричат „скачащи гени“, мобилни генетични елементи или транспозони. По време на транспониране гените, които са променили позицията си, могат да променят своята активност - това явление се нарича позиционен ефект. В резултат на позиционния ефект гените променят първоначалните си функции, което по същество води до появата на нови гени.
Транслокации- Това е преместването на участъци от хромозома или цяла хромозома към друга хромозома.
Хромозомни пренареждания
В някои случаи се получава пълно сливане на хомоложни хромозоми с образуването на двуцентромерни структури - дицентрици. В други случаи, едноцентромерна хромозома с двойно рамо се образува от две акроцентрични хромозоми. Това сливане на хромозоми се нарича Робертсонова транслокация. Робъртсъновите транслокации са често срещани при гризачи.
Последствията от хромозомните аберации варират при различните организми. В сравнително слабо организирани организми (растения, насекоми, гризачи) хромозомните пренареждания могат да доведат до появата на нови признаци, но може да не се проявят фенотипно.
При хората хромозомните пренареждания в хетерозиготно състояние намаляват плодовитостта, а в хомозиготно състояние са летални.
Механизмите на възникване на хромозомните аберации са разнообразни:
- неравномерно кръстосване между хомоложни хромозоми (възникват делеции и дупликации) и нехомоложни хромозоми (възникват транслокации);
- интрахромозомно кръстосване (възникват делеции и инверсии);
- хромозомни счупвания (появяват се различни фрагменти);
- хромозомни прекъсвания, последвани от свързване на фрагменти (възникват инверсии, транспозиции, транслокации);
- копиране на ген и прехвърляне на копие в друга част от хромозомата (възникват транспозиции).
Причините за хромозомните аберации и механизмите на тяхното възникване са различни:
- Хромозомни аберации могат да възникнат спонтанно в дългосрочно съхранявани семена или тъканни клетъчни култури без видима причина.
- Появата на хромозомни аберации се улеснява от различни химични вещества, които не са мутагени, но нарушават нормалното функциониране на клетките (йони на тежки метали, алдехиди, окислители и др.).
- Хромозомни аберации често възникват при облъчване на клетките.
В този случай се появяват както единични, така и двойни (или множествени) хромозомни прекъсвания. Единичните прекъсвания водят до появата на терминални недостатъци, двойните (множествени) прекъсвания водят до появата на всички останали видове аберации. Когато се появят прекъсвания на пресинтетичния етап, се променя цялата хромозома и се наблюдават двойни аберации; Когато се появят прекъсвания на постсинтетичния етап, само един хроматид се променя и се наблюдават единични аберации.
За идентифициране на хромозомни аберации се използват различни методи за цитогенетичен анализ.
Например анафазният анализ дава възможност да се идентифицират мостове и лагове (дицентрици и други продукти на транслокация), фрагменти (ацентрици). Метафазният и пахитеновият анализ могат да разкрият промени в хромозомната структура, линейни и кръгови фрагменти. Специално място при идентифицирането на хромозомни аберации заема анализът на гигантски политенови хромозоми, открити в слюнчените жлези на ларвите на двукрилите (комари, мухи) и в някои клетки на други организми.
Този метод се основава на нарушаване на нормалното соматично конюгиране на политенови хромозоми в хетерозиготи за хромозомни аберации; В резултат на това се образуват бримки с различна форма.
Социални бутони за Joomla
Някои мутации водят до промени в структурата на отделните хромозоми, например загуба на част от хромозома или, обратно, нейното удвояване.
1) Делеция - загуба на част от хромозома
Наблюдават се и структурни пренареждания: например част от хромозома може да се откъсне и да се премести в друга, дори нехомоложна на първата. До какво ще доведе подобно преструктуриране? Целият въпрос е кои части от хромозомите ще бъдат засегнати от мутацията. Ако такава мутация причини липсата на ензим, който е от първостепенно значение за метаболизма, тогава организмът ще умре.
Преместването на фрагмент от хромозома на място, което не е типично за него, може да промени активността на гените, които съдържа; например, "мълчаливите" гени, намирайки се в нова среда, ще започнат да работят. (Смята се, че туморните заболявания възникват точно по този начин: поради движения в хромозомата, някои „мълчаливи“ гени се озовават в онези региони, където гените в момента работят активно. В този случай не само гените, необходими на клетката, но също и чужд ген, той също започва да работи и продуктите от неговата дейност вече не се изискват от клетката).
Структурните пренареждания на хромозомите, разбира се, водят до голямо разнообразие от деформации.
Най-тежките и изразени малформации на човешкото развитие се наблюдават при така наречените геномни (да не се бъркат с генни) мутации, които се състоят от добавяне или загуба на една или повече хромозоми, както и увеличаване на броя на наборите от хромозоми.
Хромозомните мутации са мутации, които нарушават съществуващите групи на свързване или пораждат нови групи на свързване.
Това определение се отнася до начина, по който тези мутации се откриват на първо място. Според друга дефиниция хромозомните мутации са мутации, причинени от хромозомни пренареждания. Хромозомните пренареждания се предлагат в различни видове. Може би най-разпространената е рекомбинацията или кросинговърът, при която се разменят хомоложни участъци от хромозоми (фиг. 112). Други видове хромозомни пренареждания са транслокации, инверсии, делеции и дупликации.
Има различни възможности за промяна на морфологията на хромозомите.
Различават се следните КП: - Реципрочни транслокации - обмен на хромозомни участъци. — Робъртсъновите транслокации са сливането на две акроцентрични хромозоми в една хромозома с двойно рамо.
- Парацентрична инверсия - промяна в реда на гените към обратната в рамките на област, която не засяга центромера. - Перицентричната инверсия е същата, но в рамките на зоната, която включва центромера. — Вмъкването е вмъкването на допълнителен хромозомен материал в която и да е част от хромозомата. — Делеция — загубата на част от HP хромозомата води до промени в кариотипа.
| Защита на личните данни |
Не намерихте това, което търсихте? Използвайте търсачката.
Хромозомни мутации (пренареждания или аберации)- Това са промени в структурата на хромозомите, които могат да бъдат идентифицирани и изследвани под светлинен микроскоп.
Известни са различни видове пренареждания:
- липса на,или предизвикателство,- загуба на крайните участъци на хромозомата;
- изтриване- загуба на част от хромозомата в средната й част;
- дублиране -двойно или многократно повторение на гени, локализирани в определена област на хромозомата;
- инверсия- завъртане на хромозомна секция на 180 °, в резултат на което гените в тази секция са разположени в обратна последователност в сравнение с обичайната;
- транслокация- промяна в позицията на която и да е част от хромозомата в хромозомния набор. Най-често срещаният тип транслокации са реципрочни, при които регионите се обменят между две нехомоложни хромозоми. Част от хромозома може да промени позицията си без реципрочен обмен, оставайки в същата хромозома или се включва в друга.
При недостатъци, изтриванияИ дублиранеколичеството на генетичния материал се променя. Степента на фенотипна промяна зависи от това колко големи са съответните хромозомни региони и дали съдържат важни гени. Примери за недостатъци са известни в много организми, включително хора. Тежка наследствена болест - синдром на "котешки вик".(наречено на естеството на звуците, издавани от болни бебета) се причинява от хетерозиготност за дефицит на 5-та хромозома. Този синдром е придружен от тежко увреждане на растежа и умствена изостаналост. Децата с този синдром обикновено умират рано, но някои оцеляват
Геномни мутации- промяна в броя на хромозомите в генома на телесните клетки. Това явление протича в две посоки: към увеличаване на броя на цели хаплоидни набори (полиплоидия)и към загуба или включване на отделни хромозоми (анеуплоидия).
Полиплоидия- многократно увеличаване на хаплоидния набор от хромозоми. Клетките с различен брой хаплоидни набори от хромозоми се наричат триплоидни (3n), тетраплоидни (4n), хексанлоидни (6n), октаплоидни (8n) и др.
Най-често полиплоидите се образуват, когато редът на хромозомната дивергенция към клетъчните полюси е нарушен по време на мейоза или митоза. Това може да бъде причинено от физични и химични фактори. Химикали като колхицин потискат образуването на митотичното вретено в клетките, които са започнали да се делят, в резултат на което дублираните хромозоми не се разделят и клетката става тетраедрична.
За много растения т.нар полиплоидна серия.Те включват форми от 2 до 10n и повече. Например, полиплоидна серия от комплекти от 12, 24, 36, 48, 60, 72, 96, 108 и 144 хромозоми е изградена от представители на рода Solanum. Родът пшеница (Triticum) представлява серия, чиито членове имат 34, 28 и 42 хромозоми.
Полиплоидията води до промени в характеристиките на даден организъм и следователно е важен източник на вариации в еволюцията и селекцията, особено при растенията. Това се дължи на факта, че хермафродитизмът (самоопрашване), апомиксисът (партеногенезата) и вегетативното размножаване са много разпространени в растителните организми. Следователно около една трета от видовете растения, които се срещат на нашата планета, са полиплоиди, а в рязко континенталните условия на високопланинския Памир растат до 85% от полиплоидите. Почти всички културни растения също са полиплоиди, които за разлика от дивите си роднини имат по-големи цветя, плодове и семена и повече хранителни вещества се натрупват в органите за съхранение (стъбла, грудки). Полиплоидите се адаптират по-лесно към неблагоприятни условия на живот и по-лесно понасят ниски температури и засушаване. Ето защо те са широко разпространени в северните и високопланинските райони.
В основата на рязкото нарастване на продуктивността на полиплоидните форми на културните растения е явлението полимери.
анеуплоидия,или хетероплоидия,- явление, при което клетките на тялото съдържат променен брой хромозоми, който не е кратен на хаплоидния набор. Анеуплоидите възникват, когато отделните хомоложни хромозоми не се разделят или се губят по време на митоза и мейоза. В резултат на неразделяне на хромозомите по време на гаметогенезата могат да възникнат зародишни клетки с допълнителни хромозоми и след това, при последващо сливане с нормални хаплоидни гамети, те образуват зигота 2n + 1 (тризомичен)върху определена хромозома. Ако в гаметата има една хромозома по-малко, тогава последващото оплождане води до образуването на зигота 1n - 1 (монозомен)на която и да е от хромозомите. Освен това има форми 2n - 2, или нулисомика,тъй като няма двойка хомоложни хромозоми и 2n + Х,или полисомия.
Анеуплоидите се срещат в растенията и животните, както и в хората. Анеуплоидните растения имат ниска жизнеспособност и плодовитост, а при хората това явление често води до безплодие и в тези случаи не се предава по наследство. При деца, родени от майки на възраст над 38 години, вероятността от анеуплоидия се увеличава (до 2,5%). В допълнение, случаите на анеуплоидия при хора причиняват хромозомни заболявания.
При двудомните животни, както в естествени, така и в изкуствени условия, полиплоидията е изключително рядка. Това се дължи на факта, че полиплоидията, причиняваща промяна в съотношението на половите хромозоми и автозомите, води до нарушаване на конюгацията на хомоложни хромозоми и по този начин усложнява определянето на пола. В резултат на това такива форми се оказват стерилни и по-малко жизнеспособни.
Хромозомните мутации са свързани с пренареждане на хромозомите и нарушаване на тяхната структура, които възникват по време на клетъчното делене или в резултат на физически влияния. Откриването на хромозомните пренареждания даде на изследователите нов метод за анализ на генотипа и изучаване на локализацията на гените върху хромозомата. Хромозомните пренареждания играят важна роля в еволюцията на генотипа, представлявайки механизъм за генна рекомбинация както в рамките на една хромозома, така и между нехомоложни хромозоми; те осигуряват допълнителна рекомбинация в генотипа.
Въпреки че хромозомите са в състояние да възстановят структурата си, понякога такива смущения продължават и пораждат нови клетки и организми с хромозомни пренареждания, наречени хромозомни пренареждания. аберации .
От цитологична гледна точка аберациите се делят на хромозомна И хроматид . Това зависи от това кога се случват пренарежданията - преди или след хромозомната репликация. В зависимост от характера на настъпващото преструктуриране има вътре- И междухромозомни аберации (фиг. 43).
Интрахромозомните пренареждания се делят на предизвикателство , или край на недостига; изтривания – загуба на средните части на хромозомата, дублиране , или удвояване (умножаване) на част от хромозома; инверсии – промени в редуването на гените в хромозомата поради завъртане на хромозомния участък на 180°. Междухромозомните пренареждания включват транслокации - преместване на част от една хромозома към друга, която не е хомоложна.
Ориз. 43. Хромозомни пренареждания
Специално място заемат транспозициите и инсерциите - промени в локализацията на малки участъци от генетичен материал, включително един или повече гени. Транспонирането може да се случи както между нехомоложни хромозоми, така и в рамките на една и съща хромозома. Следователно транспозициите заемат междинна позиция между интра- и междухромозомните пренареждания.
Изтривания и недостатъци. Тези хромозомни пренареждания се класифицират като недостатъци. Поради недостатъци хромозомите се съкращават и физическото отсъствие на част от един от хомолозите води до хемизиготно състояние на гените, разположени в нормалния хомолог. Ако доминиращите алели на един от хомолозите на хетерозиготата са загубени, тогава се наблюдава фенотипна проява на рецесивните алели на хромозомата, които не са засегнати от аберацията. Дефицитите, които възникват в резултат на две едновременни прекъсвания в средата на хромозомата и елиминирането на вътрешна част, се наричат делеции. Прекъсванията се свързват и хромозомата става по-къса.
Цитологично, делециите могат да бъдат идентифицирани чрез появата на бримка по време на конюгацията на хомоложни хромозоми в мейозата. Делециите позволяват да се определи точната позиция на гените, които се изследват върху хромозомните карти.
Ако възникне прекъсване в едно от рамената на хромозомата и крайният участък се загуби, тогава такива недостатъци се класифицират като терминал (терминал) или дефиниция. Понякога се случват прекъсвания едновременно в две рамена на хромозома, в резултат на което двата края се елиминират. В този случай отворените краища могат да се съединят, образувайки пръстеновидна хромозома в митоза (фиг. 44).
 |
Ориз. 44. Видове хромозомна недостатъчност:
1 - терминален недостиг;
2 - два крайни дефицита, водещи до образуването на пръстен;
3 - интрахромозомна недостатъчност.
Недостигът може да бъде голям или малък. Малките делеции включват така наречените микроделеции, които могат да бъдат открити само чрез изследване на гигантски хромозоми. Микроделециите имитират генни мутации в техния фенотипен ефект, но се различават от тях само по отсъствието на ефекта на обратната мутация. Големите дефицити обикновено са смъртоносни в хомозиготно състояние, тъй като те нарушават генния баланс. Само хетерозиготи за недостатъци могат да бъдат жизнеспособни. В този случай недостатъците се проявяват като доминиращи гени.
Методът на диференциалното хромозомно оцветяване отваря големи възможности за идентифициране на делеции, дефицити и други хромозомни пренареждания. Някои багрила различно оцветяват различни области на хромозомите. Поради това хромозомите придобиват характерни напречни ивици. Този метод се използва за определяне на хромозомни пренареждания на метафазни хромозоми.
Дублиране . Това пренареждане е свързано с двукратно повторение на една и съща хромозомна секция. Има известни случаи на многократни повторения или анимации на който и да е раздел. Те се наричат още усилвания. Дублирането може да възникне в рамките на една и съща хромозома или да бъде придружено от прехвърляне на копие на част от генетичен материал към друга хромозома (транспониране). Повторенията, които възникват на една хромозома, могат да бъдат разположени тандемно ( ABCBCDE...) или обърнат ( ABCBDE…). Терминалните повторения също се разграничават, ако дублирането засяга края на хромозомата.
Много дублирания и изтривания възникват поради неравномерно пресичане. Когато подобни ДНК последователности се появят в съседни области на хромозома, конюгирането на хомолози може да се случи неправилно. Пресичането на такива неправилно конюгирани хромозомни области води до образуването на гамети с дублиране или делеция.
Цитологично хетерозиготите за дупликации се откриват подобно на хетерозиготите за делеции - по време на конюгацията на хромозомите се образува бримка. Дублиранията обикновено нямат същото отрицателно въздействие върху жизнеспособността като изтриванията и недостатъците. Подобни елементи често се повтарят в геномите на различни организми.
Дублиранията играят важна роля в еволюцията на генома, защото създават допълнителни участъци от генетичен материал, чиято функция може да бъде променена от мутация и последващ естествен подбор.
Инверсии. Инверсията е завъртане на 180° на отделни участъци от хромозомата, но броят на гените в хромозомата не се променя. Ако последователността на гените в оригиналната хромозома е обозначена А Б В Г Д, а зоната претърпя инверсия BCD, тогава на новата хромозома гените ще бъдат разположени в последователността ADCBE.
По отношение на центромера инверсиите се разделят на перицентричен , улавяйки центромера и включвайки го в обърнатата секция, и парацентричен , които не включват центромера в обърнатата област. В организми, хомозиготни за хромозомна инверсия, последователността на генното свързване се променя. Хомозиготен за инверсия ADCBEген Атясно свързан с гена д, не с IN, както в оригиналната хромозома А Б В Г Д, а ген дсвързан с IN, не с д.
Инверсиие широко разпространен път на еволюционна трансформация на генетичен материал. Например, хората и шимпанзетата се различават по броя на хромозомите: хората имат 2n=46, а шимпанзетата имат 2n=48. При хората хромозома 2 съдържа по-голямата част от материала, хомоложен на допълнителната двойка хромозоми на шимпанзето. В допълнение, разликите засягат четири хромозоми: 4-та, 5-та, 12-та и 17-та, в които са настъпили перицентрични инверсии. При хората хромозома 17 е акроцентрична, докато същата хромозома при шимпанзетата е метацентрична.
При хетерозиготите, базирани на инверсии, цитологичните препарати показват характерни примки - резултат от конюгация на променена и нормална хромозома. В този случай гените прилягат много плътно един към друг. Следователно има взаимно привличане с огромна сила между хомоложни хромозомни локуси.
Инверсиите имат еволюционно значение. Някои тясно свързани видове се характеризират с наличието на инверсии в хромозомите. Така в природата е установено, че има две раси от един и същи вид дрозофила, които не се кръстосват, а причината за безплодието е специфична инверсия във всяка от тях. Некръстосването на такива раси по същество е началото разминаване видове, тъй като мутациите, възникващи в една раса, не могат да бъдат предадени на друга; следователно тяхната еволюция трябва да поеме по различни пътища.
Транслокации. Транслокацията е реципрочен обмен на участъци от нехомоложни хромозоми. Те се отнасят до междухромозомни пренареждания, които променят групите на свързване на гените. Ако изобразим последователността на гените в оригиналните хромозоми като А Б В Г Д ЕИ KLMNO, тогава в транслокираните хромозоми генните последователности могат да бъдат, например, ABCDNOИ KLMEF. При хомозиготите за тези транслокации, в сравнение с оригиналните хромозоми, естеството на връзката се променя: гените, които не са били свързани в оригиналните хромозоми, стават свързани и обратно. Да, гени НЕсе оказват свързани с ABCDи престават да бъдат свързани с KLM. В хетерозигота за транслокация, гените, принадлежащи към различни, нехомоложни хромозоми, се наследяват като принадлежащи към една и съща група на свързване. Това се обяснява с факта, че само онези спори (гамети), които носят родителски комбинации от хромозоми, са напълно функционални.
Характер спрежение промени в транслокираните хромозоми: образува се кръстосана форма. Плътното конюгиране близо до точките на прекъсване е възпрепятствано, което води до потискане на кросинговъра в тези области.
На цитологичните препарати хетерозиготите в профаза на мейозата образуват квадриваленти, а не биваленти, както обикновено, тъй като хомоложни области се появяват на всичките четири хромозоми. Когато хиазмата се плъзне от центромерите към краищата на хромозомите, кръстът се трансформира в пръстен. Понякога хромозомите на пръстена се обръщат и образуват фигура от осем. От шестте възможни типа хаплоидни гамети само два функционират нормално - тези, които получават пълни набори от гени, характерни за оригиналните родителски форми. Останалите четири вида гамети носят дупликации и делеции и следователно, като правило, не произвеждат жизнеспособно потомство или не участват в оплождането.
Хетерозиготите за танлокации са частично стерилни, тъй като произвеждат дефектни гамети по време на процеса на мейоза. Подобно на инверсиите, транслокациите осигуряват изолация на формите и насърчават дивергенцията в рамките на един вид.
В някои случаи е възможно практическото използване на транслокации, например при копринената буба. Известно е, че добивът на копринени нишки е по-висок в мъжките пашкули. Затова в бубарството е по-изгодно да се отглеждат мъжки гъсеници. За да реши този проблем, руският генетик V.A. Струников успя експериментално да създаде мутантна линия, в която всички мъже са постоянно хетерозиготни за два неалелни рецесивни летални гена ( l 1И l 2), локализирани в различни З-мъжки хромозоми. Тези мъже са доста жизнеспособни, тъй като експресията на двата смъртоносни гена е потисната от техните доминантни алели ( L 1И L 2). Когато мъжките със смъртоносни гени, транслокирани в половите хромозоми, се кръстосват с нормални женски, само мъжките оцеляват в тяхното потомство, а женските зиготи ( WZ) умират още в яйцето, тъй като единственият З- хромозомата на тези зиготи идва от бащата и носи или един смъртоносен ген ( l 1), или друг ( l 2), а на W хромозомата липсват доминантни алели на тези гени. Получаването на мъжки гъсеници осигурява висок икономически ефект.
Транспониране и вмъкване. Транспонирането е движението на малки участъци от генетичен материал в една и съща хромозома или между различни хромозоми. Транспозициите се извършват с участието на специални мобилни или мигриращи генетични елементи. Те са два вида: инсерции - относително къси ДНК последователности, които носят информацията, необходима за собственото им транспониране, и транспозони, които освен информацията, необходима за транспониране, кодират фенотипни характеристики.
Първият мобилен (мобилен, транспониращ) генетичен елемент е описан през 1947 г. от Б. Макклинток в царевица. Тя откри миграционен локус Ds(дисоциатор), при който често се случват счупвания на хромозоми.
Самият локус Dsне предизвиква разкъсвания. Те се появяват в него само ако в генома присъства друг мигриращ елемент - ак(активатор). И двата елемента могат да бъдат загубени с честота от няколко процента в мейотичното потомство или да променят локализацията си по време на митотични деления. При което Dsдвижи се само в присъствие Асо.
Внедряване Dsв непосредствена близост до или в рамките на ген СЪС, който контролира червеното оцветяване на алейрона на семената, доведе до инактивирането на този ген и хетерозиготните семена се оказаха неоцветени. INприсъствие Асодисоциатор Dsзапочна да се движи - понякога напуска локуса СЪС. В резултат на това върху неоцветените семена се появяват цветни петна от алейрон.
Контролните елементи са клас мутации, които причиняват обезцветяване на зърната. Тези мутации бяха наречени контролни елементи, защото контролират активността на гена, отговорен за синтеза на пигмент в дадена зърнена клетка. Понякога те се връщат към дивия тип и се случва връщането към дивия тип на един мутантен ген да е придружено от появата на същата мутация в друг ген. Може да се предположи, че контролният елемент "скача" от един антоцианинов локус в друг, т.е. Макклинток описва способността на контролните елементи да се движат в целия геном.
Едва през 80-те години на ХХ век, благодарение на успехите на генното инженерство, е възможно да се изолират и изследват Асо, Dsи други мигриращи елементи в царевицата. Оказа се, че Ds- това е дефектен изтрит вариант Асо. Елементна структура Асосе оказаха типични за мигриращи елементи, които са били изследвани при бактерии, дрозофила и дрожди.
Изследването на молекулярната структура на мигриращите генетични елементи започва с откриването на много необичайни мутанти в Escherichia coli ( E. coli). Всички мутанти имат общи вмъквания с по-голяма или по-малка дължина. Те са интегрирани в различни части на генома E. coliДНК молекулите се наричат Е- елементи (от англ последователности на вмъкване– последователности на вмъкване). Открито е, че няколко различни последователности на вмъкване могат да причинят мутации в много гени. Те се различават по размер, но имат някои общи структурни характеристики. Краищата съдържат идентични или почти идентични нуклеотидни последователности, но подредени в обратен ред. Освен това, когато инсерция се вмъкне в целева ДНК, малка част от целевата ДНК последователност се повтаря близо до всеки край на инсерцията. Тази повтаряща се ДНК последователност, фланкираща инсерцията, обикновено съдържа 5 до 9 нуклеотида.
Последователностите за вмъкване са относително малки и кодират само функциите, необходими за тяхното транспониране. Вторият клас подвижни елементи, т.нар транспозони , съдържа гени, които не са свързани с транспонирането, но придават важни свойства на клетките на бактерията гостоприемник.
Транспозоните са открити за първи път, когато е установено, че някои гени за резистентност към антибиотици са свързани с фактори на инфекциозна резистентност. Индивидуалните резистентни гени могат да бъдат прехвърлени към други плазмиди и хромозоми на бактерии, поради което възниква терминът „транспозон“.
Една от полезните функции на транспозоните е, че те допринасят за включването на нови, „чужди“ гени в генома на организмите. Друга възможна функция на транспонируемите елементи може да бъде свързана с тяхната способност да причиняват различни хромозомни пренареждания, по-специално съседни делеции и инверсии. Това може да е важен механизъм за създаване на вътрешноспецифични вариации в хромозомните структури.
Ефект на позицията. Транспозициите и вмъкванията понякога активират близък ген, водят до промяна във фенотипа поради факта, че преместените гени се оказват в нова среда, а понякога и самият преместен ген се променя. Това явление се нарича ефект на позицията. Ефектът на позицията може да бъде стабилен И нестабилен , или мозайка.
Геномните мутации ще бъдат обсъдени в следващата глава.
Въпроси за обсъждане
1. Какво е модификационна променливост и какво е нейното значение?
2. Какво е спонтанен мутационен процес и какви са неговите модели?
3. Какво е процес на индуцирана мутация и какви са неговите модели?
4. Какво се разбира под честота на мутации?
5. Кои мутации се считат за директни и кои са обратни?
6. Каква е разликата между соматичните мутации и мутациите в зародишните клетки? Доминантни и рецесивни мутации?
7. Понякога точковите мутации се наричат генни мутации. Смятате ли, че това е приемливо? Защо?
8. Обяснете каква връзка съществува между мутациите и гените? Мутации и рекомбинации?
9. Когато една хромозома е счупена, точката на счупване лежи ли в гена, между гените или са възможни и двете? Опитайте се да аргументирате отговора си.
10. Каква е същността на закона за хомоложните серии на наследствената променливост? Назовете автора на този закон.
11. Обяснете понятията: дефицит, дупликация, инверсия, транслокация. Как възникват тези хромозомни аномалии?
12. Какво представляват „транспозоните“ или „скачащите гени“?
13. Обяснете същността на „ефекта на позицията на гена“.
14. Колко постоянна трябва да бъде промяната в нуклеотид, за да се счита за мутационна?
15. Заместването на кои азотни основи води до промени в типа на прехода и трансверсията?
16. Начертайте възможно най-много различни резултати от разкъсвания на хромозоми. AB/CDE/FGHJ/J, където точката обозначава центромера, а наклонените линии показват местата на три едновременни разкъсвания. Посочете кой от резултатите трябва да се появява най-често.
17. Как възникват пръстенните хромозоми?
18. Хромозома A.BCDEEDCFGима обърната област на дублиране CDE
Може също да се интересувате от:
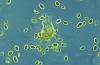
Обикновената амеба (царство Животни, подцарство Протозои) има друго име -...

Фетишизмът е разстройство на сексуалната ориентация, което се основава на сексуален...

Кожните промени при псориазис могат да бъдат локализирани във всяка част на тялото. Псориатичен...
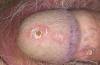
Псориазисът в интимната зона е доста често срещан както при мъжете, така и при жените. болест...

Гъбичното възпаление на влагалището е често срещано при възрастни жени и всяка поне веднъж...

